×
![]()
Corresponding Author: Valérie Maxime
Institut de Recherche Dupuy De Lôme, UMR CNRS 6027, Université Bretagne Sud, C.E.R. Yves Coppens, Campus de Tohannic, 56017 Vannes Cedex (France)
Tel. +33 297 017 169 , E-Mail valerie.maxime@univ-ubs.fr
Evidence of Coordinated and Adjustable Osmolytes Movements Following Hyposmotic Swelling in Rainbow Trout Red Blood Cells
Valérie Maxime
Institut de Recherche Dupuy De Lôme, UMR CNRS 6027, Université Bretagne Sud, C.E.R. Yves Coppens, Campus de Tohannic, Vannes, France
Introduction
The maintenance of a constant volume is a prerequisite for normal cellular life. Fish are good models for cell volume regulation studies because their aqueous habitats expose them to a wide range of steady or fluctuating osmotic conditions. Many studies demonstrate that most fish cells are able to regulate their volume following their physical swelling [1]. Notably, given their circulation through epithelial tissues, red blood cells (RBCs) can be often exposed to osmotic challenges and subsequent volume disturbances. Thus, besides the relative ease with which they are isolated, RBCs are often used as models for volume regulation studies.
In vivo cell volume increase can be induced by exposure to hyposmotic extracellular fluid or by the increase in intracellular osmolytes concentrations as a consequence of a variety of metabolic or pathologic conditions. Fish RBCs, particularly, can also transiently swell in isosmotic conditions after the stimulation of the membrane Na+/H+ exchanger by catecholamine hormones [2]. Whatever the cause, cell swelling induces an adaptive response named regulatory volume decrease (RVD) consisting of passive loss of inorganic and/or of some low molecular weight organic osmolytes, followed by osmotically-obliged water efflux leading to the restoration of cell volume. Because of its electrochemical gradient, K+ is highly involved in this process. It is tightly coupled to its counterion Cl- thanks to electroneutral cotransporters. This coupling can also take place via parallel conductive pathways [3]. Among the different classes of cell organic compounds (amino acids, polyols and methyl-amines – [4]), the β-amino acid taurine plays a primary role in RVD, due to its relatively high concentration. It can also withstand large changes in its concentration without deleterious effects on cellular function [4]. It is present in teleost’s RBCs at concentrations up to 55mM [5]. In this way, a significant hyposmotic swelling-activated taurine efflux has been demonstrated in RBCs of lamprey [6] and succeeding elasmobranchs and bone fish: skate [7], eel [8, 9], carp [10], rainbow trout [11, 12, 13], Atlantic salmon [14] and flounder [8, 15, 16]. Moreover, few works, all performed in rainbow trout [11, 12, 17, 18], describe an additional, so far unexplained, uptake of Na+ in RBCs following an hyposmotic swelling. Such an effect is rather observed after a hypertonic cell shrinkage [3]. Its clear that the RVD process based on these osmolytes effluxes has to be controlled in a well-coordinated
manner [19]. However, this aspect has received little attention. In addition, cell volume regulation is likely to induce a net transfer of acid-base equivalents across the membrane, but results are unclear. For example, a reversible and unexplained decrease in intracellular pH (pHi) of trout hepatocytes was induced by an hyposmotic stress [20]. On the contrary, a great stability of pHi during RVD process was reported in trout RBCs [11].
The first aim of this study was to estimate the respective importance of the osmotically active solutes involved in the RVD process of RBCs of rainbow trout (Oncorhynchus mykiss), with a specific attention to the apparently counter-productive Na+ uptake. This intriguing result justify the choice of the fish species, since it has already been outlined, to our knowledge, solely in rainbow trout. Then, in an attempt to clarify the effect of RVD on pHi, the latter was calculated considering the link between proton and chloride transmembrane concentrations according to the Donnan equilibrium.
Materials and Methods
Fish, samplings and experimental design
Fifty rainbow trout (Oncorhynchus mykiss L.; 800-1000 g) were obtained from a commercial hatchery (Lesneven, Brittany, France) and kept for at least one week at the laboratory, under natural photoperiod and temperature (12-15°C) conditions, in a 400 l tank supplied with flowing aerated freshwater. They were fed ad libitum with commercial dry pellets until the day before the experiment. After acclimatization, the fish were lightly-anaesthetised by immersion in tricaine methanesulfonate (Sigma – 50 mg. l-1). Whole blood samples (approximately 10 ml) were collected in heparinised syringes (10 IU/ml) by caudal puncture. Each sample allowed to perform two RVD challenges which conditions were randomly determined.
RBCs were exposed to isosmotic or one of three different hyposmotic conditions of the same level: The first one (Hypo. I) corresponds to a 50% dilution of the main component (NaCl) of the physiologic saline. The two others were made up in order to control the K+ and Cl- transmembrane concentrations. In this way, Hypo. II saline had a high K+ concentration suitable to abolish the transmembrane gradient of K+. In the Hypo. III saline, NO3- was used as a Cl- substitute to coerce Cl- to get out of the cell and to reduce its contribution to the RVD. Salines compositions are given in Table 1.
Measurements
Cells images analysis being unsuitable to the non-spherical shape of fish RBCs, their volume changes were indirectly estimated from their variations in water content according to a method allowing also the simultaneous measurement of the main ions contents [11]. The blood sample was centrifuged (2500 g for
5 min) and the plasma and buffy coat removed by aspiration. The RBCs pellet was washed 3 times in isosmotic Cortland saline [21] (ω = 295.9 mOsm.kg-1). After overnight normoxic incubation in a thermostatically controlled incubator (15°C), the cells were supposed to be not in a catecholamine-stimulated condition [10]. The sample was then adjusted to 15% haematocrit with isosmotic saline and transferred to a tonometer supplied with humidified air at 15°C. The cell suspension was hence allowed to equilibrate for at least 1 hour under slight agitation, to guarantee a steady state with respect to ions and water.
All the experiments were performed under normoxic conditions. Cells were exposed or not (control) to a moderate hyposmotic stress (ω ≈ 176 mOsm.kg-1). This was achieved, after centrifugation (2,500 g for
3 min) of the suspension, by the substitution of the isosmotic saline for one of the iso- or hyposmotic salines. Ten minutes before and then at intervals (5, 10, 20, 30, 60 and 90 min) after the hyposmotic stress, 2.5 ml aliquots of the whole suspension were drawn from the tonometer. They were immediately poured into six 400-µl nylon tubes and centrifuged at 30,000 g (Eppendorf centrifuge 5417C) for 5 min. The resulting mass of packed cells was then separated from the supernatant by slicing the tube with a scalpel blade at the top of the red cell column. Cell water content was determined in duplicate by the gravimetric method comparing the cells masses before and after their total drying for 72 hours (80°C). It was expressed as gram water per gram dry cell solids. In duplicate samples of haemolysed cells (perchloric acid 70%), intracellular K+ and Na+ contents on the one hand, and Cl- content on the other, were determined by photometry (Instrumentation Laboratory flame photometer IL 243-05), and by colorimetry (Biolabo, kit 80005), respectively. Ion
contents were expressed as µmol per gram dry cell solids.
An attempt to dissect Na+ pathways across cells membrane was performed by a pharmaceutical approach using amiloride (Sigma). It was dissolved in dimethyl sulfoxide (DMSO) and added to RBCs suspension ten minutes before and during the hyposmotic stress to yield a final concentration of 10-4M. The lack of adverse effects of DMSO was checked beforehand. Osmolalities were measured with a vapor pressure osmometer (Wescor, model 5520).
Calculations
Considering that, in rainbow trout RBCs, no solute, other than Na+, K+, Cl- and taurine significantly contribute to RVD [11] and that the net loss of osmolytes is followed by an isosmotic loss of water, the taurine contribution as a function of time after swelling was calculated from the measured net changes of water, K+, Cl- and Na+ contents. This approach is consistent with the method used by the above-mentioned authors who show that the calculated data correspond closely with the experimental ones. The hyposmotic salines containing on average 176 mmol of osmolyte per litre of water, 5.68 ml water is lost or gained for each mmol of osmolyte leaving or entering the cell. This calculation allowed also to estimate the respective participation of each inorganic osmolyte to RVD.
The intracellular pH (pHi) was calculated from the chloride distribution across the RBCs membranes according to the Donnan equilibrium theory (pHi = pHo + log [Cl-]i/[Cl-]o). The intracellular pH measurement using intracellular microelectrode or the freeze-thaw technique in fish RBCs produces results in agreement with this theory [22]. In the present study, pHo is the imposed pH of the experimental salines (Table 1) [Cl-]o
was calculated from the chloride salts composition of these solutions (132 mM and 70 mM for isosmotic and hyposmotic salines, respectively) [Cl-]i expressed also in mM, was calculated by dividing Cl- content by water content for each condition. pHi calculation is theoretically impossible for Hypo. III condition due to the lack of chlorides in the extracellular medium. Nevertheless, its trend has been estimated assuming a fictive concentration of 1 mM for this anion.
Statistical analyses
Statistical analyses were conducted using Sigmastat 12.5 (Systat Software Inc.). All data were expressed as mean values ± standard error of the means (S.E.M.). Differences were tested by a one-way ANOVA analysis. Whenever significant effects were detected, multiple comparisons were conducted using Holm-Sidak method. Differences were considered significant at p < 0.05.
Results
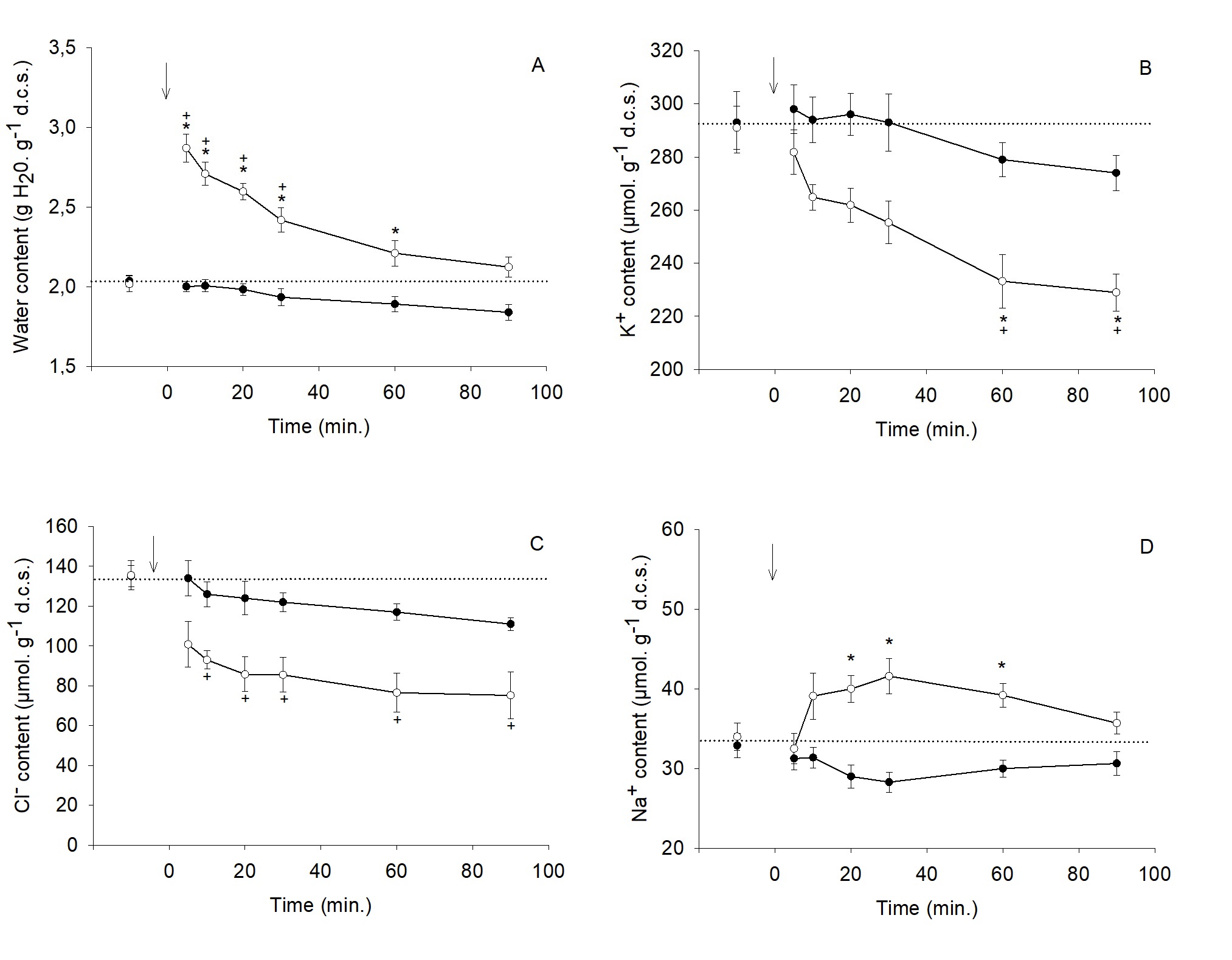
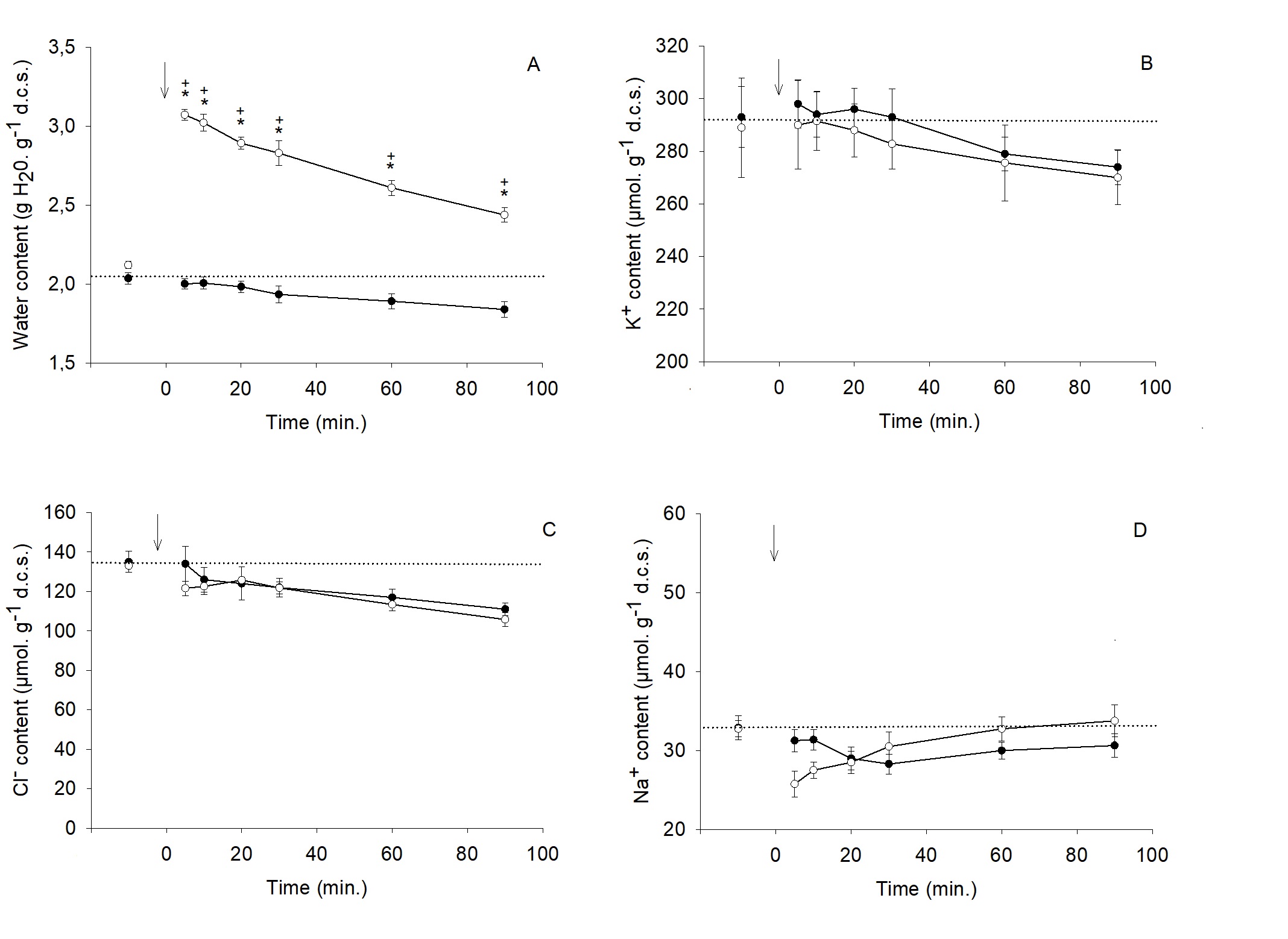
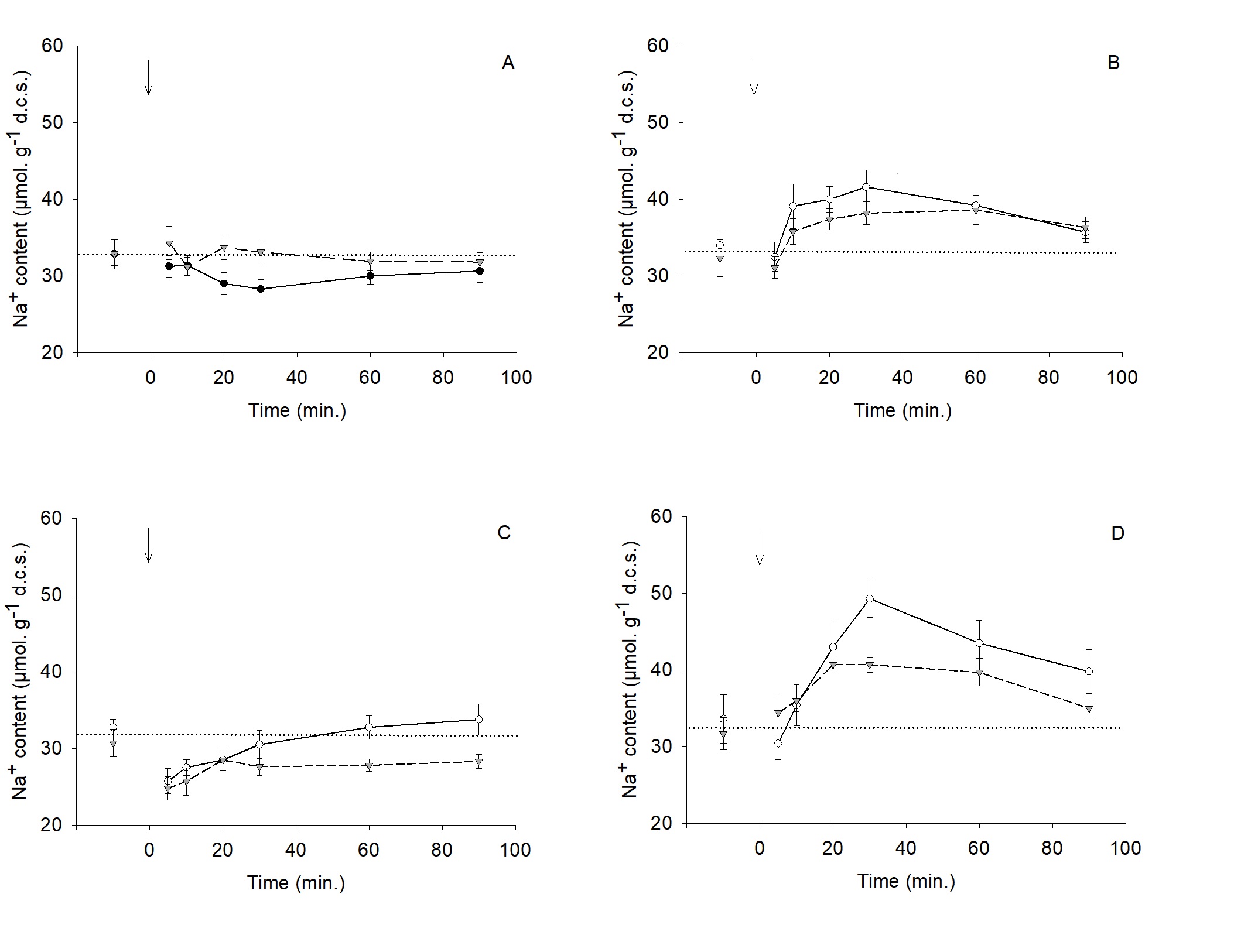
In isosmotic saline (Fig. 1-3), the mean water content of trout RBCs was 2.03 ± 0.03 g. g-1 d.c.s. and slightly decreased over the 90 min. experimental period (-11%). K+ and Cl- contents (293 and 135 µmol. g-1 d.c.s., respectively) followed the same trend (-7 and -21%, respectively). The RBCs suspension in the dilution of the physiological saline (Hypo. I condition) induced a rapid 43% increase in water content, which was nearly cleared within 90 minutes (Fig. 1A). As shown in Fig. 1B-C, this effect was concurrent with a decrease in both K+ and Cl- contents (-21% and -44%, respectively). A net Na+ influx (+ 22%) culminating after 30 minutes was also observed (Fig. 1D). According to Fig. 2A, when RBCs are in the hyposmotic saline with a high K+ concentration (Hypo. II condition), the extent of the water content increase was slightly higher than the one observed in Hypo. I condition and the restoration was incomplete (66% after 90 minutes). This condition did not bring any significant change in K+, Cl- and Na+ contents compared to isosmotic status (Fig. 2A-C). In a Cl--free saline (Hypo. III condition), RBCs fully downregulated their water content after the same magnitude increase as in
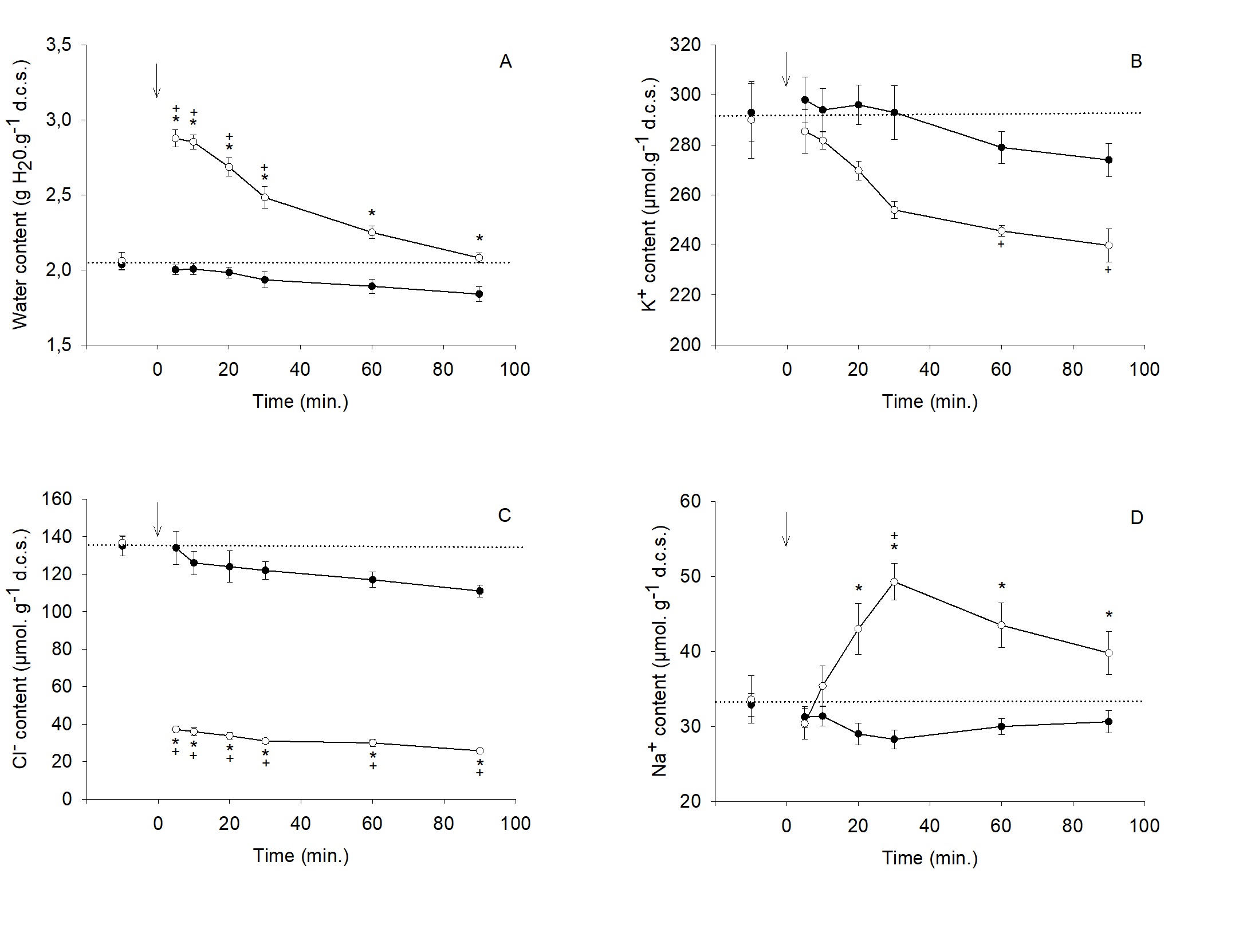
Hypo. I condition (Fig. 3A). The magnitude of the K+ content decrease was quite similar with (- 21% - Fig. 1B) or without Cl- (-17% - Fig. 3B). Fig. 3C shows that the Cl- content decreased drastically within the first five minutes due to an enlarged transmembrane gradient, to settle to a very low level (about 30 µmol. g-1 d.c.s). A significant increase in Na+ content (+ 46% at 30 min.) was also observed in this condition (Fig. 3D).
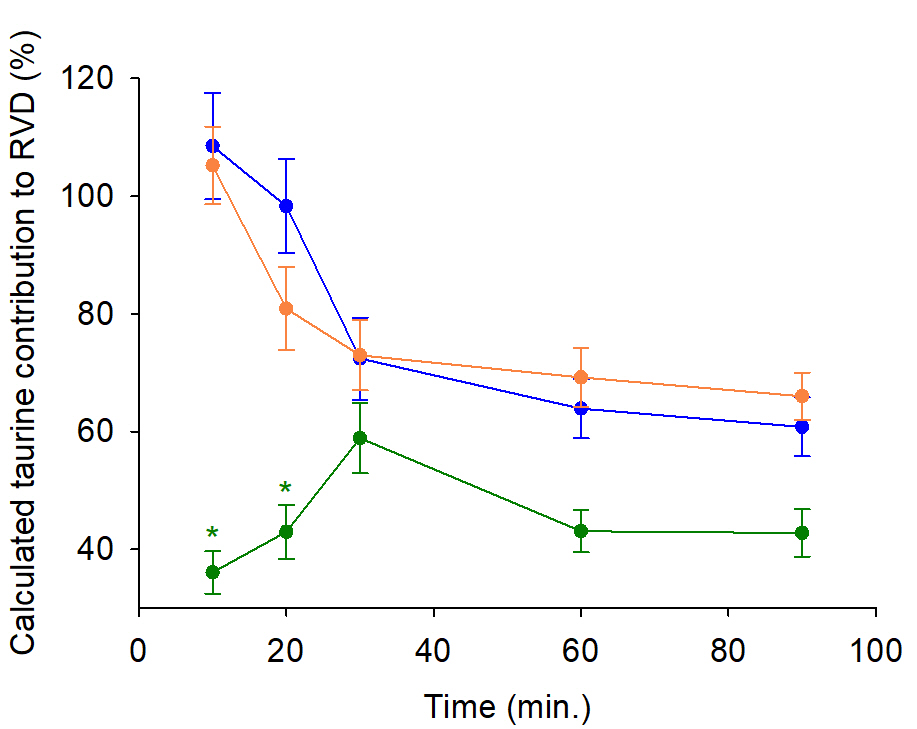
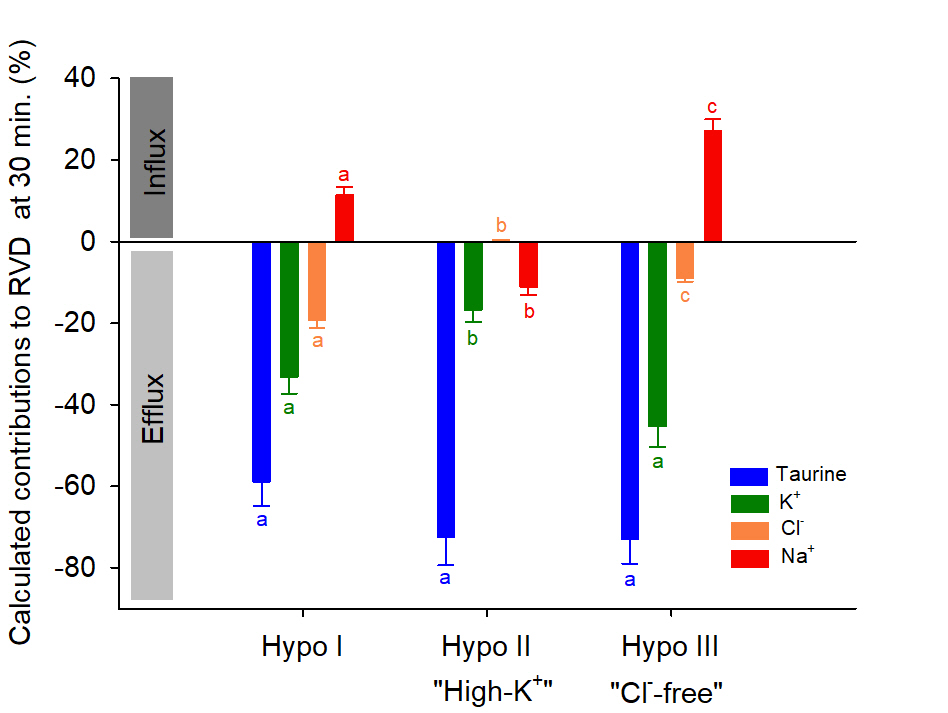
The measured losses of water, K+ and Cl-, and uptake of Na+ allowed the calculation of taurine loss and its contribution to RVD in the three hyposmotic conditions as a function of time (Fig. 4). It was estimated on average to be 45% to the RVD process in Hypo. I condition. But, it was at first close to 100%, then progressively settled down to about 69% in Hypo. II and Hypo. III conditions.
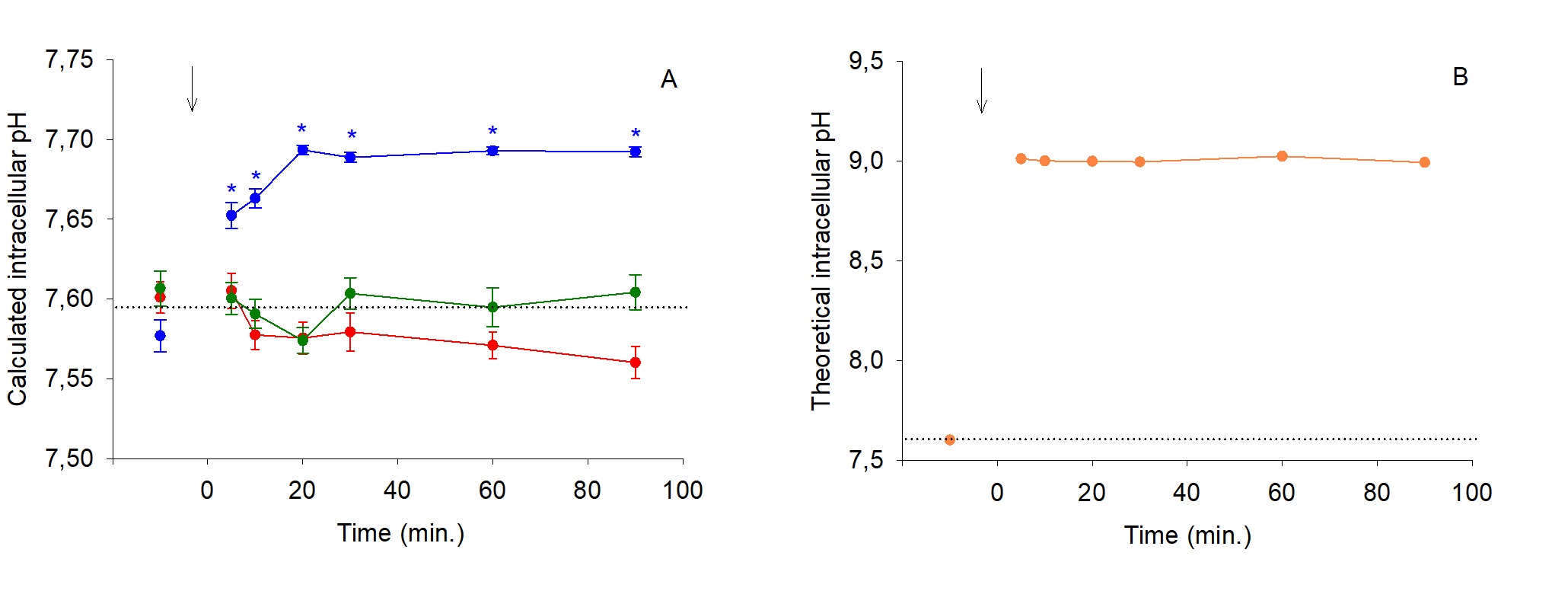
The Na+ influx was non significantly inhibited by an extracellular 10-4M application of amiloride in all hyposmotic conditions (Fig. 5A-D). In the absence of hyposmotic shock, calculated pHi values (Fig. 6A-B) are in agreement with those reported for in vitro experiments realized in same conditions [22, 23]. Fig. 6A shows a similar stability of pHi for isosmotic and Hypo. I physiological status. But, Hypo. II condition induced a significant alkalosis (ΔpHi = +0.1). Fig. 6B highlights a theoretical drastic alkalosis trend in Hypo. III condition.
Fig. 7 is an overview of the ionic and organic osmolytes calculated relative contributions to RVD after a 30 minutes RBCs exposure to hyposmotic salines. This delay corresponds to the maximal Na+ content increase. It shows the significant role played by taurine and highlights the modulations of the osmolytes fluxes resulting from the different experimental conditions.
The author thanks A. Mézergues for her technical assistance and Pr. O. Sire for his careful and critical reading of the manuscript.
Statement of Ethics
Animal procedures were in accordance with the French (articles R214-87 à R214-137 du code rural) and EU (Directive 2010/63/EU) guidelines for animal research.
The author declares that no conflict of interests exists.
| 1 Chara O, Espelt MV, Krumschabel G, Schwarzbaum PJ: Regulatory volume decrease and P receptor signalling in fish cells: mechanisms, physiology, and modelling approaches. J Exp Zool 2011;315:175-202. https://doi.org/10.1002/jez.662 |
||||
| 2 Baroin A, Garcia-Romeu F, Lamarre T, Motais R: A transient sodium-hydrogen exchange system induced by catecholamines in erythrocytes of rainbow trout (Salmo gairdneri). J Physiol 1984;356:21-31. https://doi.org/10.1113/jphysiol.1984.sp015450 |
||||
| 3 Hoffmann EK, Lambert IH, Pedersen SF: Physiology of cell volume regulation in vertebrates. Physiol Rev 2009;89:193-277. https://doi.org/10.1152/physrev.00037.2007 |
||||
| 4 Kirk K: Swelling-activated Organic Osmolyte Channels. J Membr Biol 1997;158:1-16. https://doi.org/10.1007/s002329900239 |
||||
| 5 Motais R, Guizouarn H, Borgèse F: The swelling-sensitive osmotic channel; in Bernhardt I, Ellory JC (eds): Red cell membrane transport in health and disease. Berlin/Heidelberg/New York, Springer-Verlag, 2003, pp 153-171. https://doi.org/10.1007/978-3-662-05181-8_7 |
||||
| 6 Brill SR, Musch MW, Goldstein L: Taurine efflux, band 3 and erythrocyte volume of hagfish (Myxina glutinosa) and lamprey (Petromyzon marinus). J Exp Zool 1992;264:19-25. https://doi.org/10.1002/jez.1402640104 |
||||
| 7 Davis-Amaral EM, Musch MW, Goldstein L: Chloride and taurine effluxes occur by different pathways in skate erythrocytes. Am J Physiol 1996;271:R1544-R1549. https://doi.org/10.1152/ajpregu.1996.271.6.R1544 |
||||
| 8 Fincham DA, Wolowyk MW, Young JD: Volume sensitive taurine transport in fish erythrocytes. J Membr Biol 1987;96:45-56. https://doi.org/10.1007/BF01869333 |
||||
| 9 Lewis RA, Bursell JDH, Kirk K: Anion selectivity of the swelling-activated osmolyte channel in eel erythrocytes. J Membr Biol 1996;149:103-111. https://doi.org/10.1007/s002329900011 |
||||
| 10 Jensen F: Regulatory volume decrease in carp red blood cells: mechanisms and oxygen-dependency of volume-activated potassium and amino acid transport. J Exp Biol 1995;198:155-165. https://doi.org/10.1242/jeb.198.1.155 |
||||
| 11 Guizouarn H, Motais R, Garcia-Romeu F, Borgese F: Cell volume regulation: the role of taurine loss in maintaining membrane potential and cell pH. J Physiol 2000;523:147-154. https://doi.org/10.1111/j.1469-7793.2000.t01-1-00147.x |
||||
| 12 Garcia-Romeu F, Cossins AR, Motais R: Cell volume regulation by trout erythrocytes: characteristics of the transport systems activated by hypotonic swelling. J Physiol 1991;440:547-567. https://doi.org/10.1113/jphysiol.1991.sp018724 |
||||
| 13 Kiessling K, Ellory JC, Cossins AR: The relationship between hypotonically-induced taurine and K fluxes in trout red blood cells. Pflügers Arch 2000;440:467-475. https://doi.org/10.1007/s004240000296 |
||||
| 14 Wormser C, Mason LZ, Helm EM, Light DB: Regulatory volume response following hypotonic stress in Atlantic salmon erythrocytes. Fish Physiol Biochem 2011;37:745-759. https://doi.org/10.1007/s10695-011-9474-3 |
||||
| 15 Nonnotte G, Truchot JP: Cell volume regulation by erythrocytes of the euryhaline fish, Platichthys flesus, after hyposmotic stress in bicarbonate/carbon-dioxide-buffered medium. Cell Physiol Biochem 1992;2:336-348. https://doi.org/10.1159/000154656 |
||||
| 16 Thoroed SM, Fugelli K: The Na+-independent taurine influx in flounder erythrocytes and its association with the volume regulatory taurine efflux. J Exp Biol 1994;186:245-268. https://doi.org/10.1242/jeb.186.1.245 |
||||
| 17 Godart H, Ellory JC, Motais R: Regulatory volume response of erythrocytes exposed to a gradual and slow decrease in medium osmolality. Pflugers Arch 1999;437:776-779. https://doi.org/10.1007/s004240050845 |
||||
| 18 Egée S, Lapaix F, Cossins AR, Thomas SL: The role of anion and cation channels in volume regulatory responses in trout red blood cells. Bioelectrochemistry 2000;52:133-149. https://doi.org/10.1016/S0302-4598(00)00096-9 |
||||
| 19 Wehner F, Olsen H, Tinel H, Kinne-Saffran E, Kinne RKH: Cell volume regulation: osmolytes, osmolyte transport, and signal transduction. Rev Physiol Biochem Pharmacol 2003;148:1-80. https://doi.org/10.1007/s10254-003-0009-x |
||||
| 20 Krumschnabel G, Gstir R, Manzl C, Prem C, Pafundo D, Schwarzbaum PJ: Metabolic and ionic responses of trout hepatocytes to anisosmotic exposure. J Exp Biol 2003;206:1799-1808. https://doi.org/10.1242/jeb.00370 |
||||
| 21 Wolf K: Physiological salines for fresh-water teleosts. Prog Fish Cult 1963;25:135-140. https://doi.org/10.1577/1548-8659(1963)25[135:PSFFT]2.0.CO;2 |
||||
| 22 Thomas S, Egée S: Fish red blood cells: characteristics and physiological role of the membrane ion transporters. Comp Biochem Physiol A 1998;119:79-86. https://doi.org/10.1016/S1095-6433(97)00404-2 |
||||
| 23 Guizouarn H, Motais R: Swelling activation of transport pathways in erythrocytes: effects of Cl-, ionic strength, and volume changes. Am J Physiol 1999;276:C210-C220. https://doi.org/10.1152/ajpcell.1999.276.1.C210 |
||||
| 24 National Center for Biotechnology Information (2021): PubChem Annotation Record for Taurine, Source: Hazardous Substances Data Bank (HSDB) [cited 2020, August 12]. URL: https://pubchem.ncbi.nlm.nih.gov/source/hsdb/8167. | ||||
| 25 Shennan DB: Swelling-induced taurine Transport: relationship with chloride channels, anion-exchangers and other swelling-activated transport pathways. Cell Physiol Biochem 2008;21:15-28. https://doi.org/10.1159/000113743 |
||||
| 26 Guizouarn H, Gabillat N, Motais R, Borgèse F: Multiple transport functions of a red blood cell anion exchanger, tAE1: its role in cell volume regulation. J Physiol 2001;535:497-506. https://doi.org/10.1111/j.1469-7793.2001.t01-1-00497.x |
||||
| 27 Huxtable RJ: Physiological actions of taurine. Physiol Rev 1992;72:101-163. https://doi.org/10.1152/physrev.1992.72.1.101 |
||||
| 28 Alvarez de la Rosa D, Canessa CM, Fyfe GK, Zhang P: Structure and regulation of amiloride sensitive sodium channels. Annu Rev Physiol 2000;62:573-594. https://doi.org/10.1146/annurev.physiol.62.1.573 |
||||
| 29 Harris C, Fliegel: Amiloride and the Na+/H+ exchanger protein: mechanism and significance of the Na+/H+ exchanger. Int J Mol Med 1999;3:315-321. https://doi.org/10.3892/ijmm.3.3.315 |
||||
| 30 Chesney RW, Gusowski N, Dabbagh S, Theissen M, Padilla M, Diehl A: Factors affecting the transport of β-amino acids in rat renal brush-border membrane vesicles. The role of external chloride. Biochim Biophys Acta 1985;812:702-712. https://doi.org/10.1016/0005-2736(85)90264-0 |
||||
| 31 Zelikovic I, Chesney RW: Ionic requirements for amino acid transport. Am J Kidney Dis 1989;14:313-316. https://doi.org/10.1016/S0272-6386(89)80211-2 |
||||
| 32 Jones DP, Miller LA, Dowling C, Chesney RW: Regulation of taurine transporter activity in LLC-PK1 cells: Role of protein synthesis and protein kinase C activation. J Am Soc Nephrol 1991;2:1021-1029. https://doi.org/10.1681/ASN.V251021 |
||||
| 33 Malapert M, Guizouarn H, Fievet B, Jahns R, Garcia-Romeu F, Motais R, Borgèse F: Regulation of Na+/H+ antiporter in trout red blood cells. J Exp Biol 1997;200:353-360. https://doi.org/10.1242/jeb.200.2.353 |
||||
| 34 Chen XJ, Eaton DC, Jain L: Alveolar epithelial ion and fluid transport β-adrenergic regulation of amiloride-sensitive lung sodium channels. Am J Physiol Lung Cell Mol Physiol 2002;282:L609-L620. https://doi.org/10.1152/ajplung.00356.2001 |
||||