×
![]()
Corresponding Author: Marco G. Alves
Institute of Biomedical Sciences Abel Salazar, Department of Anatomy and Unit for Multidisciplinary
Research in Biomedicine (UMIB),
School of Medicine and Biomedical Sciences (ICBAS), University of Porto, R. Jorge de Viterbo Ferreira 228, Porto, 4050-313 (Portugal)
Tel. +351 96 7245248 , E-Mail alvesmarc@gmail.com
Hyperoside Supplementation in Preservation Media Surpasses Vitamin C Protection Against Oxidative Stress-Induced Damages in Human Spermatozoa
Mafalda V. Moreiraa,b Sara C. Pereirab Bárbara Guerra-Carvalhob,c,d David F. Carragetab Soraia Pintoe Alberto Barrose,f,g Branca M. Silvah Pedro F. Oliveirac Marco G. Alvesb
aFaculty of Sciences, University of Porto, Porto, Portugal, bDepartment of Anatomy and UMIB - Unit for Multidisciplinary Research in Biomedicine, ICBAS - School of Medicine and Biomedical Sciences, University of Porto, Porto, Portugal, cQOPNA & LAQV, Department of Chemistry, University of Aveiro, Aveiro, Portugal, dDepartment of Life Sciences, Faculty of Sciences and Technology, University of Coimbra, Coimbra, Portugal, eCentre for Reproductive Genetics Professor Alberto Barros, Porto, Portugal, fDepartment of Genetics, Faculty of Medicine, University of Porto, Porto, Portugal, gi3S – Instituto de Investigação e Inovação em Saúde, University of Porto, Porto, Portugal, hUniversity of Beira Interior, Covilhã, Portugal
Introduction
The decline in male fertility is an emerging problem in modern societies. The increasing trend for couples to have children at a later age, along with unhealthy lifestyles, and detrimental environmental factors are reducing the general fertility rates. According to the World Health Organization (WHO), infertility can be defined as a disease from the reproductive system that prevents a couple from conceiving, after 12 months of regular and unprotected sexual activity. This health issue affects approximately 15% of couples worldwide and is thought that the male factor is associated with around half of these cases [1]. Multiple causes affect male reproductive potential. However, about 30-50% of cases of male infertility have unexplained causes and are classified as idiopathic. Recently, it has been proposed that idiopathic infertility could be explained by the presence of high levels of reactive oxygen species (ROS) [2]. In fact, evidence suggests that ROS-mediated damage contributes to about 30-80% of male infertility cases [3].
In normal physiological conditions, reactive oxygen species (ROS) and antioxidants balance each other, which is essential for the maintenance of several molecular signalling pathways. However, when ROS exceeds the antioxidant capacity, the cell turns into a state of oxidative stress (OS). Prolonged exposure to OS is extremely prejudicial to cellular systems, promoting the disruption of signalling pathways, affecting essential metabolic processes, resulting in the degradation of cellular machinery [4]. The male reproductive system is very sensitive to OS. Spermatozoa are cells especially vulnerable to ROS-attack due to the abundance of polyunsaturated fatty acids (PUFAs) in their plasma membrane. PUFAs are essential for plasma membrane fluidity, which is essential for zona pellucida binding and the process of fertilization [5]. Furthermore, spermatozoa are known to carry a very limited amount of cytoplasm, which results in a lack of antioxidant defences, and inefficiency in repairing oxidative damage [6]. ROS are known for playing a dual role in biological systems, having prejudicial and beneficial effects. Evidence suggests that certain levels of ROS are required for physiological sperm functions, including sperm capacitation, hyperactivation, acrosomal reaction, and sperm-oocyte fusion [7-9]. On the other hand, in spermatozoa, the overproduction of ROS results in lipid peroxidation (LPO) of membranes [10], protein modification [11] DNA damage [12], and increased apoptosis. These pathological events have been negatively correlated with sperm motility [13], sperm morphology [14], and male fertility capacity [15].
Couples affected by infertility often choose to pursue parenthood through the help of assisted reproduction technology (ART). The implementation of ART allowed several infertile couples to achieve a successful pregnancy. However, the success rate of ART is still considered low, being at best 50.4 %, in < 35-year-old women with 3 good quality embryos, according to the 2011 study of Vernon, et al. [16]. ART is also known to increase the exposure of gametes and embryos to ROS. In the case of male gametes, the removal of seminal plasma, as well as sperm processing techniques (centrifugation, light exposure, temperature) causes an increase in ROS production and promotes a reduction in sperm quality [17], decreasing (even more) the chances of achieving a successful pregnancy [18]. To prevent the oxidative damage of spermatozoa during ART, the sperm preparation medium is often supplemented with antioxidants.
Hyperoside (quercetin 3-O -β-D-galactoside) (HYP) is a flavonol glycoside, commonly found in plants of the genera Hypericum [19] and Crataegus [20]. It is a product of quercetin glycosylation of the hydroxyl group on C-3, with a galactoside group [21]. HYP extraction and isolation are difficult, due to their low concentrations, and their synthesis is costly. Nevertheless, this phytochemical has gained popularity due to its anti-cancer [22], anti-inflammatory [23], anti-thrombotic [24], and anti-depressant [25] pharmacological properties. Recent studies have also explored the antioxidant effects of HYP. HYP was reported to possess cytoprotective properties against OS in lung fibroblast cells [26]. Also, it was reported that HYP protects granulosa cells against hydrogen peroxide (H2O2)-induced cell apoptosis and OS, revealing potential benefits on female reproductive capacity [27]. Biagi M. and colleagues reported that Castanea sativa Miller leaf extract, particularly rich in HYP, showed scavenging properties against OS in human sperm and showed being capable to protect sperm membranes and the acrosomes [28]. All these findings suggest that HYP can be used as a potential antioxidant in disorders associated with oxidative damage. On the other hand, VC (L-ascorbic acid) is a water-soluble vitamin known for its antioxidant properties. This micronutrient is a reducing agent and acts like an electron donor that reacts with substrates such as H2O2, generating ascorbyl radicals, a relatively stable free radical [29]. VC cannot be synthesized by human metabolism, meaning that its uptake comes straight from food [30]. Its concentration in the seminal fluid can be 10 times higher than in the serum, with values ranging between 200-700 µM [31-35]. VC is commonly prescribed in oral antioxidant therapies for infertile men (usually in combination with other compounds), and it is reported to decrease sperm DNA fragmentation and improve sperm-quality parameters [36-39]. In vitro studies did also report that the supplementation of sperm medium with VC was able to decrease negative effects of vitrification on sperm parameters and chromatin quality [40], and improve DNA integrity [41, 42].
To our knowledge, no studies have been performed regarding the potential of HYP supplementation of the sperm preservation medium used during ART. Thus, we propose to test and compare the efficacy of sperm preservation media supplementation with HYP and VC in the prevention of spermatozoa-oxidative damage. To test the sperm-protective potential of these compounds, we supplemented Ham’s F12 medium with VC (600 µM) and HYP (100 and 500 µM) and maintained spermatozoa for one hour at 37°C. Sperm motility and vitality, were evaluated before and after treatment. Subsequently, protein and lipid oxidative damage, DNA fragmentation, and mitochondrial membrane potential (MMP) of spermatozoa were assessed. The total antioxidant capacity (TAC) of the treatment media was also evaluated, and the spermatozoa metabolic profile was investigated.
Materials and Methods
Patient characterization and study design
Human sperm samples were collected from Centro de Genética da Reprodução Professor Alberto Barros, located in Porto, Portugal, after approval by the Joint Ethics Committee CHUP/ICBAS (2021/CE/P002[P342/CETI/ICBAS]) during November 2020 and June 2021. The semen samples were provided by male patients of reproductive age undergoing fertility assessment, after 2-4 days of abstinence. Semen samples were obtained by masturbation and placed in sterile tubes. Patients with inflammatory diseases, cancer, or other health conditions that may severely affect their fertility potential were discarded. Also, patients were asked to provide information on the consumption of tobacco, alcohol, and other substances that could interfere with the experiment. All patients included in this study signed informed written consent. Sperm samples were characterized according to the WHO guidelines for laboratory examination and processing of human sperm [43]. Only samples having the following normal semen parameters, according to the WHO-criteria, were used in this project: sperm concentration ≥ 15 million/mL, total sperm motility ≥ 40%, and sperm vitality ≥ 58%. The fresh semen samples were centrifuged at 500 g for 5 minutes at room temperature. The seminal fluid was discarded, and the pellet was washed with 1 mL of phosphate-buffered saline (PBS) solution. After another centrifugation at 500 g for 5 minutes, the supernatant was discarded, and the pellet was resuspended in Ham F12 medium.
To test if HYP (HWI group, Rülzheim, Germany) had a cytotoxic effect in human spermatozoa, sperm samples (10 million sperm/condition) were incubated with increasing concentrations of HYP (0, 5, 50, 100, and 500 µM), for 1h at 37°C, 5% CO2. Sperm vitality and sperm motility were measured before and after the experiment. The concentrations of 100 and 500 µM of HYP (1h of incubation time) were selected for further experiments. To test the sperm-protective potential of HYP and VC (Sigma-Aldrich, St. Louis, Missouri, USA), 10 million spermatozoa were incubated for 1h at 37°C, 5% CO2, in the following conditions: 1) Ham F12 medium; 2) Ham F12 medium + 600 µM VC; 3) Ham F12 medium + 100 µM HYP; 4) Ham F12 medium + 500 µM HYP; 5) Ham F12 medium + 300 µM H2O2; 6) Ham F12 medium + 600 µM VC + 300 µM H2O2; 7) Ham F12 medium + 100 µM HYP + 300 µM H2O2; 8) Ham F12 medium + 500 µM HYP + 300 µM H2O2. H2O2 (LabChem, Lisbon, Portugal) was used as a positive control for OS, as described in [44, 45]. VC concentration of 600 µM was chosen according to [31-35]. Sperm vitality and sperm motility were assessed before and after the experiment according to the WHO guidelines [43]. Afterwards, samples were centrifuged at 500 g , 37°C. Sperm pellets and post -treatment media were stored at -80°C until used in further analysis.
Oxidative stress evaluation
Sperm pellets were resuspended in 10 µL of 1% SDS lysis buffer (1 µL for every 106 cells). The samples were vortex and left at room temperature for 60 min to promote the lysis reaction. Subsequently, lysates were centrifuged at 14000 g for 20 minutes, at room temperature. The cell debris pellet was discarded. Protein quantification was performed using the Pierce Bicinchoninic acid protein assay (Thermo Fisher Scientific, Massachusetts, USA). Levels of lipid peroxidation, protein carbonylation, and protein nitration were evaluated by immunoblotting. For lipid peroxidation and protein nitration levels detection, 5 µg of protein was diluted in PBS to a final volume of 100 µL and transferred to polyvinylidene difluoride (PVDF) membranes through a slot-blot system. For protein carbonylation evaluation, we used the protocol described in [46]. In resume, 5 µg of protein was diluted in PBS to a final volume of 20 µL. To each sample was added 20 µL of 12% SDS solution for protein denaturation and carbonyl groups exposer. Subsequently, 40 µL of 20 mM 2,4-dinitrophenylhydrazine (DNPH) in 10% trifluoroacetic acid (TFA) was added, and the reaction was allowed to occur for 30 min at room temperature, in the dark. To stop the reaction between carbonyl groups and DNPH, 30 µL of 2M Tris with 18% β-mercaptoethanol solution was added. 2.4 µL of the derivatized samples were diluted in 107.6 µL of PBS and transferred to PVDF membranes through a slot-blot system.
Membranes were then blocked with a blocking solution [5% bovine serum albumin (BSA) in Tris-Buffered Saline, 0.1% Tween 20 (TBS-T)] for 1h30 and incubated overnight with the correspondent primary antibody. For lipid peroxidation evaluation, the primary antibody used was anti-4-hydroxynonenal (4-HNE) (1:1000 diluted in 1% BSA TBS-T; AB5605, EMD Millipore, Temecula, CA, USA). For protein nitration evaluation, the primary antibody used was anti-3-nitrotyrosine (NT) (1:1000 diluted in 1% BSA TBS-T; 9691S, Cell Signaling Technology, Danvers, MA, USA). For protein carbonylation evaluation, the primary antibody used was anti-dinitrophenol (DNP) (1:5000 diluted in 1% BSA TBS-T; D9656, Sigma-Aldrich, St. Louis, Missouri, USA). Membranes were washed with TBS-T and incubated with an appropriate secondary antibody for 1h30 [Anti-Goat 1:5000 diluted in 1% BSA TBS-T (A4187) or Anti-Rabbit 1:5000 diluted in 1% BSA TBS-T (A3687), both from Sigma-Aldrich, St. Louis, Missouri, USA]. Immuno-reactive proteins were detected through reaction with an ECF substrate (GE Healthcare, Buckinghamshire, UK), and analyzed by ChemiDocTM MP Imaging system (Bio-Rad, California, USA). Total protein was assessed and quantified by Ponceau staining. Results were normalized to total protein.
Post-treatment media total antioxidant capacity
The TAC of the post-treatment media samples was measured by Ferric Reducing Antioxidant Power (FRAP) assay as described in [47]. FRAP reagent was freshly prepared by mixing 300 mM acetate buffer (pH 3.6), 10 mM 2,4,6-tripyridyl-S-triazine (TPTZ) in 40 mM HCl and 20 mM FeCl3·6H2O in the ratio of 10:1:1.
Briefly, in a 96-well plate, 6 μL of post-treatment media samples were added to 180 µL of FRAP reagent. Triplicates were made for each sample. Distilled water (dH2O) was used as negative control and VC (1000 µM) was used as an antioxidant standard. Both controls were treated in the same way as media samples. The absorbance was read at 593 nm, immediately after the addition of the FRAP reagent (0 min) and after 40 min. Between measurements, the plate was kept in the dark, at room temperature. The antioxidant potential of the samples was determined against VC (1000 µM) standard and corrected using the absorbance value of dH2O absorbance. FRAP value (µmol antioxidant potential/L) was calculated using the formula described in [47].
Sperm DNA fragmentation evaluation
The evaluation of sperm DNA fragmentation was performed by the terminal deoxynucleotidyl transferase dUTP Nick End Labeling (TUNEL) assay, using in-situ Cell Death Detection Kit (Fluorescein) from Roche Diagnostics GmbH (Mannheim, Germany). Spermatozoa (10 million) were collected and treated with HYP, and VC as previously described. A smear of each sample was performed on adhesive slides. The slides were subsequently fixed with 4% paraformaldehyde, for 1h at room temperature and then rinsed with PBS. For cell permeabilization, slides were immersed in a solution of 0.1% sodium citrate containing 0.1% Triton X, for 2 min at 4°C. After two washes with PBS for 5 min, 50 µL of the TUNEL reaction mix [5 µL of terminal deoxynucleotidyl transferase (TdT) and 45 µL of labeled deoxyuridine triphosphate nucleotides (dUTPs) mixture] was added to each slide. A coverslip was placed to prevent evaporation. The slides were incubated in a humidified chamber at 37°C for 1h, in darkness. The slides were then washed four times with PBS (2 min each). Subsequently, coverslips were mounted using mounting medium VECTASHELD (30 Ingold Road, Burlingame, CA 94010, USA). The results were visualized in a Nikon Eclipse E400 microscope equipped with a Y-FL epi-fluorescence attachment and HB-10103AF Super high-pressure mercury lamp power supply (Nikon, Shinagawa, Tokyo, Japan), coupled with a Nikon NIS Elements Image Software. 200 spermatozoa were counted on each slide. The total number of spermatozoa was quantified under blue fluorescence (corresponding to the DAPI staining of the nucleus). Spermatozoa presenting DNA fragmentation also present green fluorescence (corresponding to the dUTPs inserted in the DNA breaks by TdT). The percentage of DNA fragmentation was calculated.
Mitochondrial membrane potential evaluation
The MMP of spermatozoa was measured by using the lipophilic probe 5-5′,6-6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolcarbocyanine iodide (JC-1) (T3168, Invitrogen™, Carlsbad, CA, USA), which can selectively penetrate mitochondria [45]. In spermatozoa with damaged mitochondrial membrane, the JC-1 remains in the cytoplasm, in a monomeric form. Meanwhile, in spermatozoa with normal mitochondrial membranes occur the accumulation of JC-1 in the membrane, promoting the formation of JC-1 aggregates. Briefly, 1 million spermatozoa were collected after treatment, washed with PBS (500 g , 5 min) and incubated with JC-1 (1 µg/mL in PBS) for 30 min at 37°C. As a positive control, spermatozoa were incubated with a 20% DMSO solution to promote the increase of mitochondrial membrane potential. After the incubation period, spermatozoa were washed with PBS (500 g , 5 min) two times. Pellets were resuspended in 750 µL of PBS and transferred to 96-well plates (performed in triplicate). The fluorescence of the JC-1 monomers (ex 485/530 nm; excitation/emission) and JC-1 aggregates (535/590 nm; excitation/emission) were assessed by a Synergy™ H1 multi-mode microplate reader (BioTek, Winooski, VT, USA). The ratio between JC-1 aggregates/monomers was calculated and used as an MMP marker.
Post-treatment media metabolite analysis
Metabolites present in the post-treatment media were assessed by proton nuclear magnetic resonance (1H-NMR) and spectra analysis. 1D 1H-NMR spectra were acquired on a 500MHz Bruker Avance III HD spectrometer equipped with a 5-mm TXI probe, at 298K. To each post -treatment medium (180 µL) it was added 45 µL of sodium fumarate (final concentration = 2 mM), for a final sample volume of 225 µL. Sodium fumarate was used as an internal reference for metabolite quantification in the media. The quantified metabolites were the following (multiplicity, chemical shift(ppm)): pyruvate (singlet, 2.37), acetate (singlet, 1.90), lactate (doublet, 1.32), alanine (doublet, 1.47), and malate (double doublet, 2.65). The spectra were manually phased, and baseline corrected. NUTS-ProTM NMR software (Acorn NMR, Inc, Fremont, CA, USA) was used to integrate the chosen metabolite peaks. The concentration of the metabolites is expressed as nmol/106 sperm cells.
Statistical Analysis
All data presented are expressed as whisker boxes (median, 25th to 75th percentiles ± minimum and maximum values). Variations between groups regarding sperm total motility, sperm vitality, OS biomarkers, DNA fragmentation, MMP, and metabolite quantification were evaluated by one-way ANOVA followed by the correction for multiple comparisons by controlling the False Discovery Rate (FDR). The two-stage step-up method of Benjamini, Krieger, and Yekutieli was the test used to assess multiple comparisons. Fold variation to the 0 µM group was calculated for OS biomarkers quantification, DNA fragmentation evaluation, and MMP assessment. The differences between the groups in the cytotoxic test and the TAC media assessment were performed using one-way ANOVA, followed by Tukey’s post hoc test for multiple comparisons. Values of P < 0.05 were considered statistically different. The statistical analysis of this work was performed by GraphPad Prism 8 software (GraphPad Software Inc., San Diego, CA, USA).
Results
Supplementation of sperm preservation media with hyperoside has no cytotoxic effects on human sperm
After treating human spermatozoa with increasing doses of HYP (in µM: 0, 5, 50, 100, and 500), for 1h at 37°C, cytotoxicity was evaluated as in [48, 49], through assessment of sperm motility and vitality. Fig. 1 represents sperm total motility and sperm vitality, after preservation in media supplemented with HYP. Regarding sperm motility, we could not find any significant differences between the HYP treated groups (in µM: 5, 50, 100, and 500), and the control group (0 µM) (Fig. 1A). Similarly, we could not find any significant differences in spermatozoa vitality after preservation in media supplemented with HYP at any concentration, and the control group (Fig. 1B). No cytotoxic effects were found in sperm samples preserved in media supplemented with any of the tested HYP concentrations. The concentrations of 100 and 500 µM of HYP were selected for further testing.
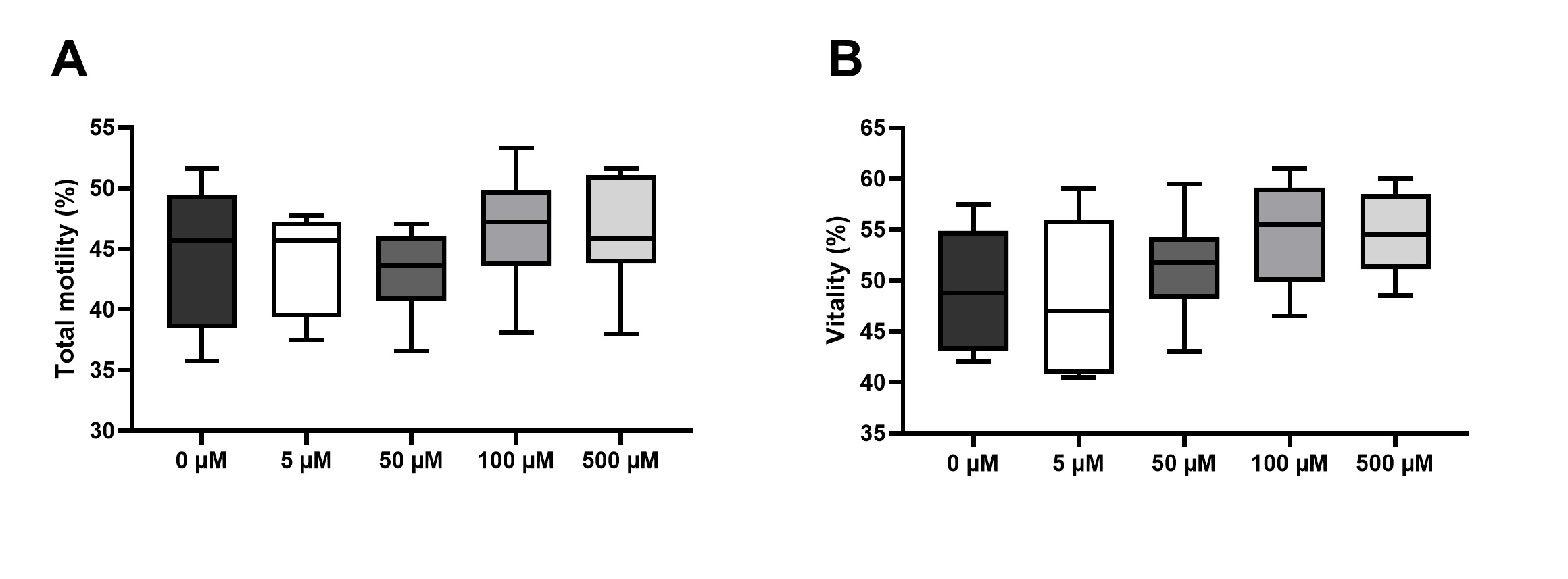
Supplementation of sperm preservation media with vitamin C and hyperoside preserves sperm motility under oxidative stress conditions
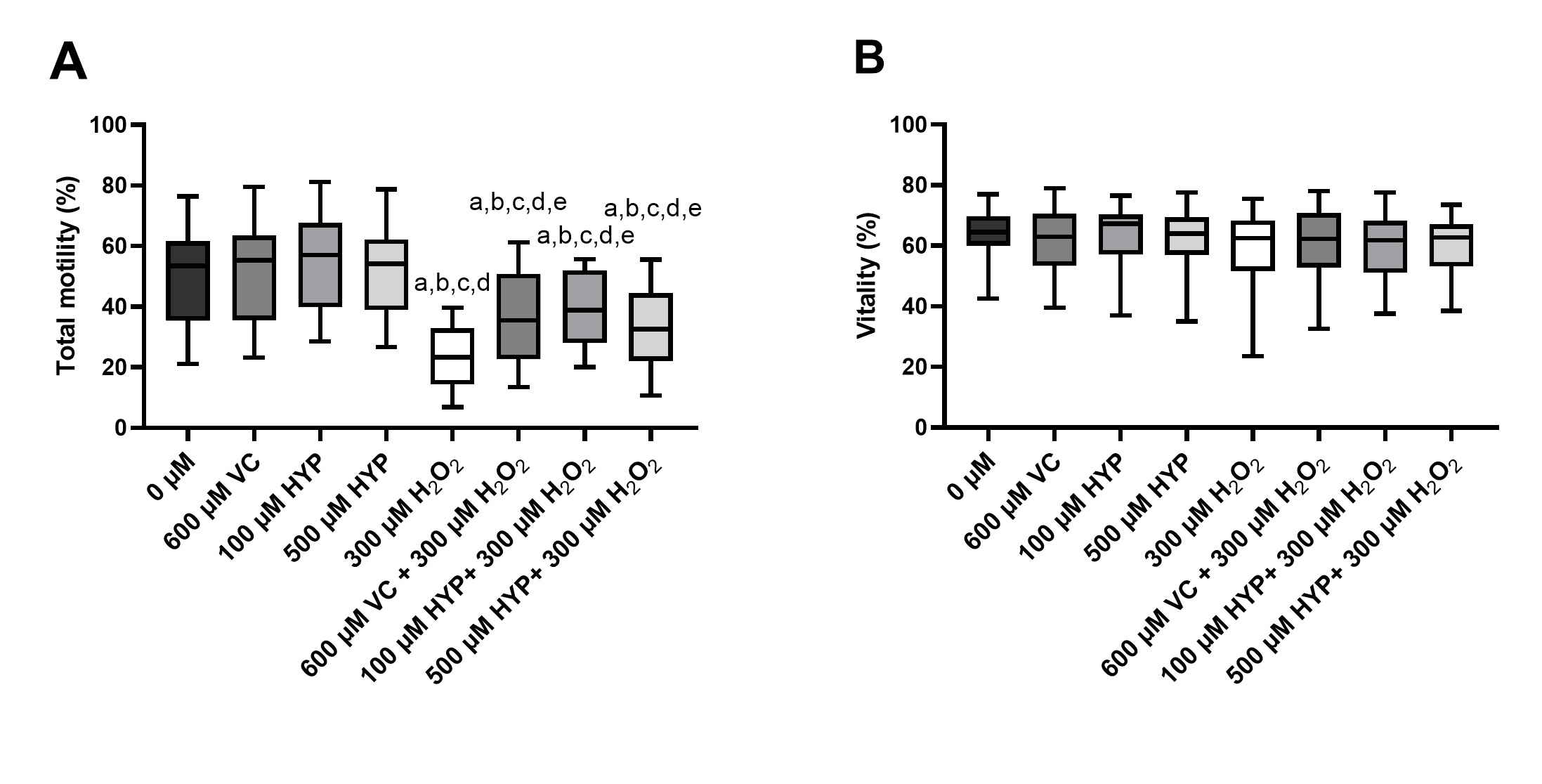
To test the cytoprotective effects of VC and HYP supplementation in human spermatozoa the following concentration were selected: 600 µM for VC [31-35]; 100 and 500 µM for HYP (after considering the previous cytotoxicity test results), 1h at 37°C. H2O2 (300 µM) was used as an OS-inducer in spermatozoa. Fig. 2 represents spermatozoa total motility and spermatozoa vitality, after preservation in media supplemented with VC and HYP, in the presence and absence of H2O2. We found no differences regarding spermatozoa total motility after preservation in media supplemented with VC or HYP, in comparison with the control group (0 µM), illustrating that supplementation of media with these antioxidants does not affect spermatozoa motility (Fig. 2A). However, spermatozoa motility decreased in the group exposed to H2O2 (300 µM) (22 ± 10 %) in comparison to the control group (50 ± 15 %) and those preserved in media supplemented with VC and HYP (100 and 500 µM) (52 ± 16 %; 55 ± 15 % and 51 ± 14 %, respectively) suggesting that spermatozoa are under stress (Fig. 2A). The spermatozoa motility of the group preserved in the media supplemented with VC and treated with H2O2 (36 ± 15 %) was higher than in the group treated with H2O2 alone suggesting a cytoprotective effect of VC supplementation against the damage promoted by H2O2. Nevertheless, the spermatozoa motility of this group was decreased in comparison to the control group and the only-antioxidant groups. The motility of the groups preserved in media supplemented with HYP (100 and 500 µM) and treated with H2O2 (39 ± 12 % and 35 ± 13 %, respectively) was higher than the motility of the group treated only with H2O2. Still, the spermatozoa motility of those groups decreased when compared to the control group and the only-antioxidant groups. These results suggest a protective effect of the antioxidants VC and HYP under OS conditions.
Regarding spermatozoa vitality, none of the preservation media supplemented with VC and HYP (100 and 500 µM) with or without H2O2 promoted a decrease of spermatozoa vitality after 1h treatment (Fig. 2B). These results suggest that, although the concentration of 300 µM of H2O2 can induce a decrease in sperm motility, this treatment does not affect sperm vitality.
Supplementation of sperm preservation media with hyperoside increases the total antioxidant capacity of the media
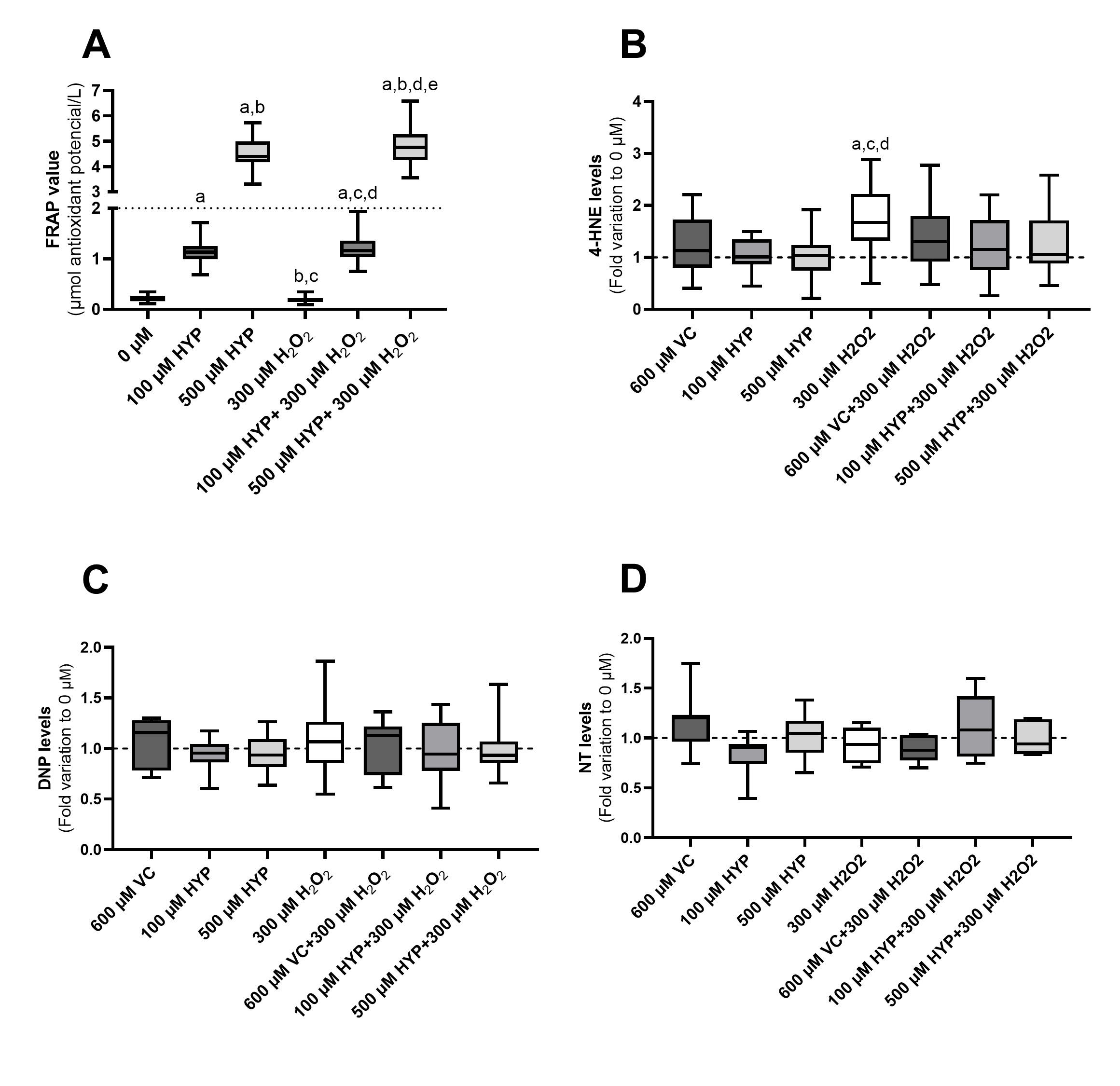
After preserving spermatozoa with media supplemented with HYP (100 and 500 µM) in the presence and absence of H2O2 for 1h at 37°C, TAC of the media was measured by FRAP. Fig. 3A represents the TAC levels of the media, herein measured by FRAP levels. VC was used as a standard for antioxidant capacity, and, because of that, TAC of the sperm preservation media supplemented with VC (600 µM) was not assessed. Nevertheless, VC (1000 µM) was used as a standard value of TAC (2 µmol antioxidant capacity/L). The media containing 500 µM of HYP (in the presence and absence of H2O2) presented the highest TAC levels (4.78 ± 0.73 and 4.54 ± 0.58 µmol antioxidant capacity/L, respectively). Also, the TAC value of the spermatozoa preservation media supplemented with 500 µM of HYP was higher than the standard TAC level of VC (1000 µM). The media containing 100 µM of HYP (in the presence and absence of H2O2) also presented higher TAC levels (1.22 ± 0.31 and 1.15 ± 0.24 µmol antioxidant capacity/L, respectively) in comparison to the control (0.21 ± 0.06 µmol antioxidant capacity/L) and only-H2O2 group (0.18 ± 0.06 µmol antioxidant capacity/L). Although the TAC of these media was lower than the ones containing 500 µM of HYP. Additionally, the TAC of the spermatozoa preservation media supplemented with 100 µM of HYP was lower than the standard TAC level of VC (1000 µM). The medium containing H2O2 (300 µM) had the lowest TAC from all the media tested. However, no difference was found regarding the TAC from this group and the TAC from the control group (0 µM). Notwithstanding, the FRAP does not react with H2O2, which can justify the absence of differences concerning the groups with the presence of H2O2 [50]. The FRAP assay measures the ability of an antioxidant compound to reduce ferric ions (Fe3+) to ferrous ions (Fe2+) and this reaction is linked to a colour change (absorbance at 593 nm) [47].
Supplementation of sperm preservation media with vitamin C and hyperoside appears to protect human sperm against lipid peroxidation under oxidative stress conditions
Elevated levels of H2O2 are known to promote OS and cause several damages to spermatozoa biomolecules, such as lipids and proteins. OS biomarkers were quantified to evaluate the oxidative damage promoted by H2O2 in human sperm and to further test the cytoprotective effects of sperm preservation media supplementation with VC and HYP. Fig. 3 (panels B, C, and D) represents the levels of lipid peroxidation, protein carbonylation, and protein nitration found in human sperm after supplementation of sperm preservation media with VC and HYP in the presence and absence of H2O2, 1h, 37°C.
Lipid peroxidation was assessed by the quantification of 4-HNE levels (Fig. 3B). Lipid peroxidation levels of the groups preserved in media supplemented with VC and HYP (100 and 500 µM) (1.25 ± 0.54; 1.04 ± 0.31 and 1.04 ± 0.45 fold, respectively) and the control group (represented as a dashed line, treatment with no HYP, no VC, and no H2O2) were similar, after 1h treatment. Meanwhile, in the group treated with only H2O2 (300 µM) (1.71 ± 0.70 fold) the spermatozoa lipid peroxidation levels increased when compared to the control group and those preserved in media supplemented with HYP (100 and 500 µM) groups. The peroxidation levels of spermatozoa preserved in media supplemented with VC and HYP (100 and 500 µM), in the presence of H2O2 (1.39 ± 0.61; 1.23 ± 0.58 and 1.30 ± 0.57 fold, respectively) were not significantly different from the levels detected in spermatozoa from the group treated with H2O2 alone (Fig. 3B). Notwithstanding, spermatozoa peroxidation levels in those groups tend to be lower in comparison to the levels detected in spermatozoa subjected only to H2O2, suggesting that cytoprotective action of VC and HYP may be occurring to some extent.
The DNP was used to assess protein carbonylation levels induced by our treatments in spermatozoa (Fig. 3C). No statistical differences were found regarding the carbonylation levels of spermatozoa preserved in media supplemented with VC and HYP (100 and 500 µM) in the presence and absence of H2O2 in comparison to the control group. Further, no statistical difference was found regarding spermatozoa carbonylation levels after treatment with H2O2 alone, and the control group (Fig. 3C). Similar results were found regarding spermatozoa protein nitration levels, evaluated by the quantification of NT (Fig. 3D). No statistical differences were found regarding the nitration levels of spermatozoa preserved in media supplemented with VC and HYP (100 and 500 µM) in the presence and absence of H2O2 in comparison to the control group. Similarly, no statistical differences were found regarding the nitration levels of the groups treated with H2O2 (300 µM), and the control group (Fig. 3D).
Supplementation of sperm preservation media with vitamin C and hyperoside prevents DNA fragmentation in human spermatozoa under oxidative stress conditions
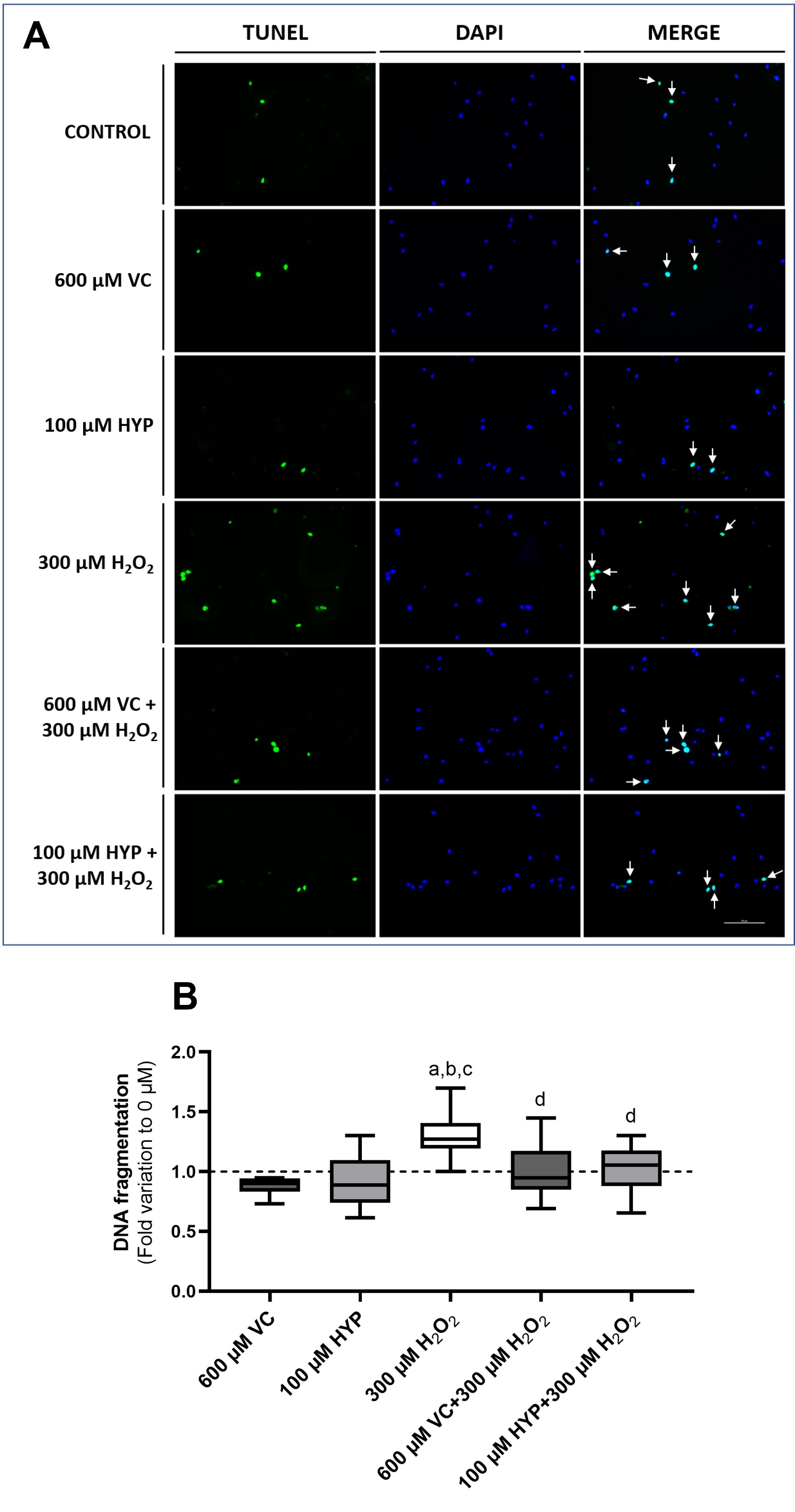
One of the consequences of a prolonged OS situation is DNA damage [12], and increased apoptosis. In spermatozoa, DNA damage is known to severely decrease the chances of achieving a successful pregnancy and increase the risk of abortion. To test if supplementation of sperm media with VC and HYP was able to prevent DNA fragmentation induced by OS we performed a TUNEL assay after preservation in the experimental conditions. Fig. 4A represents the result of the TUNEL assay visualized by fluorescence. Spermatozoa with DNA fragmentation present a green fluorescence corresponding to the dUTPs inserted in the DNA breaks. The nuclei were stained with DAPI, which has a blue fluorescence. The merge of the green/blue channels allows the detection of the percentage of sperm DNA fragmentation for each condition. Fig. 4B represents the percentage of DNA fragmentation detected in human spermatozoa after 1h incubation in media supplemented with VC and HYP (100 µM) in the presence and absence of H2O2. Our results showed no differences regarding sperm DNA fragmentation between the control group (represented as a dashed line, treatment with no HYP, no VC, and no H2O2), the VC (600 µM) (0.88±0.08-fold) and the HYP (100 µM) (0.91±0.22-fold) supplemented groups. This indicates that the supplementation with VC (600 µM) and HYP (100 µM) does not induce DNA fragmentation per se. Spermatozoa treated with H2O2 (300 µM) had higher levels of DNA fragmentation by a 1.30 ± 0.19 fold variation to the control group. Furthermore, we found that spermatozoa supplemented with VC and HYP (100 µM) in the presence of H2O2 (1.02 ± 0.23 and 1.02 ± 0.20 fold, respectively) exhibited lower levels of DNA fragmentation compared to the group treated only with H2O2. In addition, the DNA fragmentation levels of these groups did not differ from the control and antioxidants-only groups. These results may suggest that a cytoprotective effect against DNA damage could be promoted by the supplementation of VC and HYP in sperm preservation media.
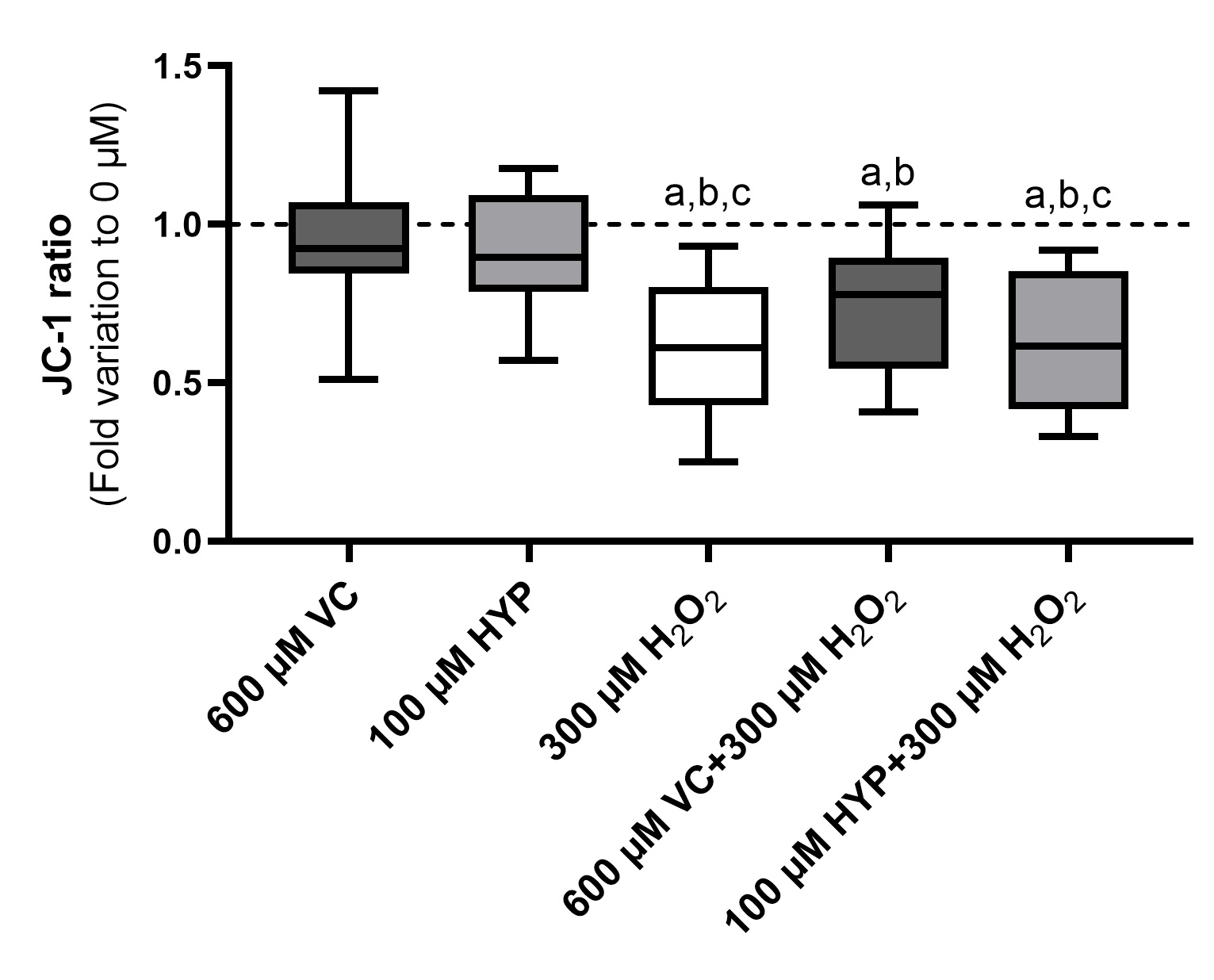
Supplementation of sperm preservation media with vitamin C and hyperoside cannot prevent alteration in the sperm mitochondrial membrane under oxidative stress conditions
MMP is a strong indicator of mitochondrial functionality and can be used as a measure of electron transport chain (ETC) activity. Mitochondria are fundamental to producing cellular energy. In spermatozoa, several studies are reporting a positive correlation between MMP and motility. To test if media supplementation with VC and HYP can ensure mitochondrial activity, under conditions of OS, we used the JC-1 probe. Fig. 5 represents the ratio of JC-1-aggregates/monomers in human spermatozoa, after 1h preservation in media supplemented with VC (600 µM) and HYP (100 µM) in the presence and absence of H2O2. Our results show no differences between the groups preserved in media supplemented with VC (600 µM) (0.97 ± 0.23 fold) and HYP (100 µM) (0.92 ± 0.19 fold) and the control group (represented as a dashed line, treatment with no HYP, no VC, and no H2O2), meaning that no MMP alterations were promoted by the antioxidant treatment, per se. A decrease in the MMP of spermatozoa can be observed in the group treated with H2O2 (300 µM) in comparison with the control group and with those preserved in media supplemented with antioxidants. However, no differences were found regarding spermatozoa MMP between the groups preserved in media supplemented with VC and HYP (100 µM), in the presence of H2O2, (0.72 ± 0.20 and 0.64 ± 0.21 fold, respectively), and the group treated with H2O2-only (0.59 ± 0.22 fold). Also, following sperm motility results (Fig. 2A), the MMP of the groups preserved in media supplemented with antioxidants treated with H2O2 was decreased when compared to MMP of spermatozoa from the control and antioxidants-only groups. This suggests that the presence of VC (600 µM) and HYP (100 µM) in the sperm media is not able to prevent the decrease in the sperm MMP promoted by H2O2.
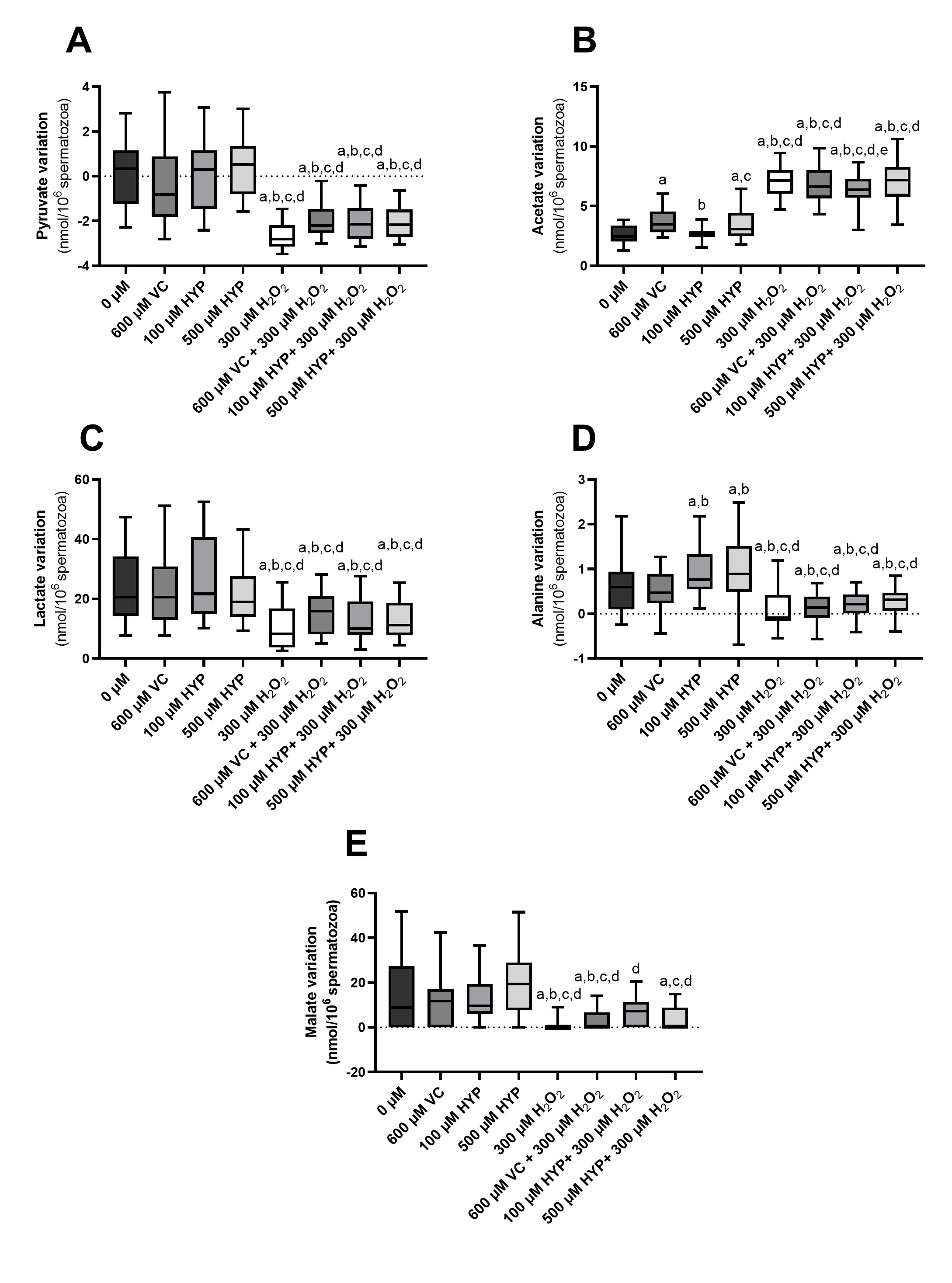
Supplementation of sperm preservation media with vitamin C and hyperoside cannot prevent alterations in the metabolism of spermatozoa under oxidative stress conditions
Metabolism is a network of biochemical reactions that ensure cell function. The study of spermatozoa metabolism can enlighten concerning what is going inside the cell. The effects of VC (600 µM) and HYP (100 and 500 µM) supplementation, in the presence and absence of H2O2, on spermatozoa metabolism were assessed by 1H-NMR spectra analysis of the media. Fig. 6 represents the main metabolites identified in the studied groups: pyruvate (Fig. 6A); acetate (Fig. 6B); lactate (Fig. 6C); alanine (Fig. 6D) and malate (Fig. 6E).
Concerning pyruvate variation, no differences were observed in pyruvate variation in the media, between the control (0.09 ± 1.58 nmol/106 cells) and the ones supplemented with VC (600 µM), and HYP (100 and 500 µM) (-0.23 ± 1.92; 0.06 ± 1.58 and 0.65 ± 1.48 nmol/106 cells, respectively) (Fig. 6A). The group treated with H2O2 presented the lowest levels of pyruvate (-2.65±0.57 nmol/106 cells). The media supplemented with VC (600 µM), and HYP (100 and 500 µM), in the presence of H2O2, (-2.00 ± 0.83; -2.11 ± 0.81 and -2.08 ± 0.74 nmol/106 cells, respectively) presented similar pyruvate levels to the H2O2-only group, which were significantly lower when compared to the control group and the antioxidants-only groups (Fig. 6A).
Regarding acetate media levels, we found that media supplementation with VC (600 µM) (3.72 ± 1.09 nmol/106 cells) and HYP (500 µM) (3.49 ± 1.27 nmol/106 cells), presented an increase in acetate production by spermatozoa compared to the control group (2.65 ± 0.76 nmol/106 cells). However, no differences were found between the acetate levels of the HYP (100 µM) group (2.66 ± 0.56 nmol/106 cells) and the control group. On the other hand, we observed higher levels of acetate in the medium of spermatozoa treated only with H2O2 (7.11 ± 1.25 nmol/106 cells) (Fig. 6B). The levels of acetate in spermatozoa from VC (600 µM) plus H2O2 group (6.80 ± 1.52 nmol/106 cells) and the HYP (500 µM) plus H2O2 group (7.04 ± 1.59 nmol/106 cells) were similar to the acetate levels of the H2O2-only group. Nevertheless, we also ascertain that acetate levels in the media of spermatozoa preserved in media supplemented with HYP (100 µM) plus H2O2 (6.35 ± 1.51 nmol/106 cells) were lower than in the only- H2O2 group (Fig. 6B).
Concerning lactate media levels, no differences were found regarding the lactate levels of the control group (24.02 ± 12.43 nmol/106 cells), the ones supplemented with VC (600 µM), and HYP (100 and 500 µM) (22.36 ± 11.85; 26.11 ± 14.05 and 21.67 ± 10.10 nmol/106 cells, respectively). Lactate production by spermatozoa was decreased in the group treated with only H2O2 (10.79 ± 7.23 nmol/106 cells) in comparison with control and the antioxidant-only groups. In addition, the levels of lactate of the media supplemented with VC (600 µM) and the HYP (100 and 500 µM), in the presence of H2O2 (15.28 ± 7.07; 13.12 ± 7.17 and 13.14 ± 6.46 nmol/106 cells, respectively) were similar to the lactate levels of the H2O2-only group (Fig. 6C) and lower when compared to the control group and the antioxidants-only groups.
Interestingly, the media of spermatozoa preserved with HYP (100 and 500 µM) presented higher levels of alanine (0.93 ± 0.60 and 1.00 ± 0.77 nmol/106 cells, respectively) when compared with the control group (0.61 ± 0.65 nmol/106 cells), and the one supplemented with VC (600 µM) (0.53 ± 0.44 nmol/106 cells). As seen in other metabolites, the media with only H2O2 supplementation (0.12 ± 0.48 nmol/106 cells), and the media supplemented with VC (600 µM) and HYP (100 and 500 µM), in the presence of H2O2 (0.13 ± 0.31; 0.20 ± 0.31 and 0.25 ± 0.34 nmol/106 cells, respectively) presented decreased alanine levels when compared to the control and antioxidants-only groups (Fig. 6D).
Concerning malate variation, no differences were found in media malate levels between the control group (14.36 ± 17.04 nmol/106 cells) and the ones supplemented with VC (600 µM), and HYP (100 and 500 µM) (11.63 ± 11.39; 12.95 ± 11.56 and 20.00 ± 15.63 nmol/106 cells). The H2O2-only group presented the lowest malate levels (1.25 ± 2.76 nmol/106 cells) in comparison to the control and the antioxidant-only groups. Decreased malate levels were also in media supplemented with VC (600 µM) and HYP (100 and 500 µM), in the presence of H2O2, (2.71 ± 4.66; 7.21 ± 6.02 and 4.33 ± 5.46 nmol/106 cells, respectively) (Fig. 6E).
Author Contributions
MVM participated in the conception, and design of the article, from the acquisition of data to bibliographic search, analysis, writing, illustration, data interpretation, and critical discussion. SCP participated in the conception, and design of this article, acquisition of data, bibliographic search, analysis, writing, data interpretation, and critical discussion. BGC participated in the acquisition of 1H-NMR spectra, spectra analysis, data interpretation, and critical discussion. DFC participated in the acquisition of 1H-NMR spectra and spectra analysis. SP provided the clinical support for this work, collecting samples and clinical data, and participating in the data analysis/ interpretation. AB provided a medical perspective to the discussion of the article. BMS, MGA, and PFO contributed to article conception and design, writing, bibliographic enrichment, data interpretation, and critical discussion. All authors approved the final version for submission.
Funding
This work was supported by Fundação para a Ciência e a Tecnologia - FCT to Sara C. Pereira (2021.05487.BD); David F. Carrageta (SFRH/BD/136779/2018); Marco G. Alves (IFCT2015 and PTDC/MEC-AND/28691/2017); LAQV-REQUIMTE (UIDB/50006/2020); UMIB (UIDB/00215/2020, and UIDP/00215/2020); ITR - Laboratory for Integrative and Translational Research in Population Health (LA/P/0064/2020). Pedro F. Oliveira was funded by national funds through FCT – Fundação para a Ciência e a Tecnologia, I.P., under the Scientific Employment Stimulus - Institutional Call - reference CEECINST/00026/2018.
Statement of Ethics
This study was approved by the Joint Ethics Committee of Centro Hospitalar Universitário do Porto (CHUP)/Institute of Biomedical Sciences Abel Salazar CHUP/ICBAS (2021/CE/P002[P342/CETI/ICBAS]). All procedures were performed in accordance with the local, national, and European ethical guidelines. All patients participated in this study voluntarily. They were informed about the purpose of the study and signed informed written consent.
The authors have no conflicts of interest to declare.
| 1 Agarwal A, Mulgund A, Hamada A, Chyatte MR: A unique view on male infertility around the globe. Reprod Biol Endocrinol 2015;13:1-9. https://doi.org/10.1186/s12958-015-0032-1 |
||||
| 2 Alahmar AT: Role of Oxidative Stress in Male Infertility: An Updated Review. J Hum Reprod Sci 2019;12:4-18. https://doi.org/10.4103/jhrs.JHRS_150_18 |
||||
| 3 Agarwal A, Parekh N, Panner Selvam MK, Henkel R, Shah R, Homa ST, Ramasamy R, Ko E, Tremellen K, Esteves S, Majzoub A, Alvarez JG, Gardner DK, Jayasena CN, Ramsay JW, Cho CL, Saleh R, Sakkas D, Hotaling JM, Lundy SD, et al.: Male Oxidative Stress Infertility (MOSI): Proposed Terminology and Clinical Practice Guidelines for Management of Idiopathic Male Infertility. World J Mens Health 2019;37:296-312. https://doi.org/10.5534/wjmh.190055 |
||||
| 4 Sies H: Oxidative stress: a concept in redox biology and medicine. Redox Biol 2015;4:180-183. https://doi.org/10.1016/j.redox.2015.01.002 |
||||
| 5 Bisht S, Faiq M, Tolahunase M, Dada R: Oxidative stress and male infertility. Nat Rev Urol 2017;14:470-485. https://doi.org/10.1038/nrurol.2017.69 |
||||
| 6 Barati E, Nikzad H, Karimian M: Oxidative stress and male infertility: current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell Mol Life Sci 2020;77:93-113. https://doi.org/10.1007/s00018-019-03253-8 |
||||
| 7 Griveau JF, Lannou DL: Reactive oxygen species and human spermatozoa: physiology and pathology. Int J Androl 1997;20:61-69. https://doi.org/10.1046/j.1365-2605.1997.00044.x |
||||
| 8 de Lamirande E, Cagnon C: Human sperm hyperactivation and capacitation as parts of an oxidative process. Free Radic Biol Med 1993;14:157-166. https://doi.org/10.1016/0891-5849(93)90006-G |
||||
| 9 Aitken RJ, Irvine DS, Wu FC: Prospective analysis of sperm-oocyte fusion and reactive oxygen species generation as criteria for the diagnosis of infertility. Am J Obstet Gynecol 1991;164:542-551. https://doi.org/10.1016/S0002-9378(11)80017-7 |
||||
| 10 Griveau JF, Dumont E, Renard P, Callegari JP, Le Lannou D: Reactive oxygen species, lipid peroxidation and enzymatic defence systems in human spermatozoa. J Reprod Fertil 1995;103:17-26. https://doi.org/10.1530/jrf.0.1030017 |
||||
| 11 O'Flaherty C, Matsushita-Fournier D: Reactive oxygen species and protein modifications in spermatozoa. Biol Reprod 2017;97:577-585. https://doi.org/10.1093/biolre/iox104 |
||||
| 12 Villani P, Eleuteri P, Grollino MG, Rescia M, Altavista P, Spanò M, Pacchierotti F, Cordelli E: Sperm DNA fragmentation induced by DNAse I and hydrogen peroxide: an in vitro comparative study among different mammalian species. Reproduction (Cambridge, England) 2010;140:445-452. https://doi.org/10.1530/REP-10-0176 |
||||
| 13 Kuroda S, Yumura Y, Mori K, Yasuda K, Takeshima T, Kawahara T, Miyoshi Y, Uemura H, Iwasaki A, Takashima K, Ikeda M, Kondo Y: Negative correlation between the presence of reactive oxygen species and Sperm Motility Index in whole semen samples of infertile males. Rev Int Androl 2017;15:84-89. https://doi.org/10.1016/j.androl.2016.08.002 |
||||
| 14 Aziz N, Saleh RA, Sharma RK, Lewis-Jones I, Esfandiari N, Thomas AJ, Jr., Agarwal A: Novel association between sperm reactive oxygen species production, sperm morphological defects, and the sperm deformity index. Fertil Steril 2004;81:349-354. https://doi.org/10.1016/j.fertnstert.2003.06.026 |
||||
| 15 Yumura Y, Iwasaki A, Saito K, Ogawa T, Hirokawa M: Effect of reactive oxygen species in semen on the pregnancy of infertile couples. Int J Urol 2009;16:202-207. https://doi.org/10.1111/j.1442-2042.2008.02213.x |
||||
| 16 Vernon M, Stern JE, Ball GD, Wininger D, Mayer J, Racowsky C: Utility of the national embryo morphology data collection by the Society for Assisted Reproductive Technologies (SART): correlation between day-3 morphology grade and live-birth outcome. Fertil Steril 2011;95:2761-2763. https://doi.org/10.1016/j.fertnstert.2011.02.008 |
||||
| 17 Agarwal A, Said TM, Bedaiwy MA, Banerjee J, Alvarez JG: Oxidative stress in an assisted reproductive techniques setting. Fertil Steril 2006;86:503-512. https://doi.org/10.1016/j.fertnstert.2006.02.088 |
||||
| 18 Gupta S, Sekhon L, Kim Y, Agarwal A: The role of oxidative stress and antioxidants in assisted reproduction. Curr Women's Health Rev 2010;6:227-238. https://doi.org/10.2174/157340410792007046 |
||||
| 19 Zou Y, Lu Y, Wei D: Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro . J Agric Food Chem 2004;52:5032-5039. https://doi.org/10.1021/jf049571r |
||||
| 20 Orhan I, Özçelik B, Kartal M, Özdeveci B, Duman H: HPLC quantification of vitexine-2 ″-O-rhamnoside and hyperoside in three Crataegus species and their antimicrobial and antiviral activities. Chromatographia 2007;66:153-157. https://doi.org/10.1365/s10337-007-0283-x |
||||
| 21 Pei J, Chen A, Zhao L, Cao F, Ding G, Xiao W: One-Pot Synthesis of Hyperoside by a Three-Enzyme Cascade Using a UDP-Galactose Regeneration System. J Agric Food Chem 2017;65:6042-6048. https://doi.org/10.1021/acs.jafc.7b02320 |
||||
| 22 Liu YH, Liu GH, Mei JJ, Wang J: The preventive effects of hyperoside on lung cancer in vitro by inducing apoptosis and inhibiting proliferation through Caspase-3 and P53 signaling pathway. Biomed Pharmacother 2016;83:381-391. https://doi.org/10.1016/j.biopha.2016.06.035 |
||||
| 23 Ku SK, Zhou W, Lee W, Han MS, Na M, Bae JS: Anti-inflammatory effects of hyperoside in human endothelial cells and in mice. Inflammation 2015;38:784-799. https://doi.org/10.1007/s10753-014-9989-8 |
||||
| 24 Ku SK, Kim TH, Lee S, Kim SM, Bae JS: Antithrombotic and profibrinolytic activities of isorhamnetin-3-O-galactoside and hyperoside. Food Chem Toxicol 2013;53:197-204. https://doi.org/10.1016/j.fct.2012.11.040 |
||||
| 25 Zheng M, Liu C, Pan F, Shi D, Zhang Y: Antidepressant-like effect of hyperoside isolated from Apocynum venetum leaves: possible cellular mechanisms. Phytomedicine 2012;19:145-149. https://doi.org/10.1016/j.phymed.2011.06.029 |
||||
| 26 Piao MJ, Kang KA, Zhang R, Ko DO, Wang ZH, You HJ, Kim HS, Kim JS, Kang SS, Hyun JW: Hyperoside prevents oxidative damage induced by hydrogen peroxide in lung fibroblast cells via an antioxidant effect. Biochimica et biophysica acta General subjects 2008;1780:1448-1457. https://doi.org/10.1016/j.bbagen.2008.07.012 |
||||
| 27 Wang X, Fan G, Wei F, Bu Y, Huang W: Hyperoside protects rat ovarian granulosa cells against hydrogen peroxide-induced injury by sonic hedgehog signaling pathway. Chem Biol Interact 2019;310:108759. https://doi.org/10.1016/j.cbi.2019.108759 |
||||
| 28 Biagi M, Noto D, Corsini M, Baini G, Cerretani D, Cappellucci G, Moretti E: Antioxidant effect of the Castanea sativa Mill. leaf extract on oxidative stress induced upon human spermatozoa. Oxid Med Cell Longev 2019;2019:8926075. https://doi.org/10.1155/2019/8926075 |
||||
| 29 Pehlivan F: Vitamin C: An Antioxidant Agent; in Hamza AH (eds): Vitamin C. IntechOpen, 2017, pp 25-33. https://doi.org/10.5772/intechopen.69660 |
||||
| 30 Lykkesfeldt J, Michels AJ, Frei B: Vitamin C. Adv Nutr 2014;5:16-18. https://doi.org/10.3945/an.113.005157 |
||||
| 31 Colagar AH, Marzony ET: Ascorbic Acid in human seminal plasma: determination and its relationship to sperm quality. J Clin Biochem Nutr 2009;45:144-149. https://doi.org/10.3164/jcbn.08-251 |
||||
| 32 Fraga CG, Motchnik PA, Shigenaga MK, Helbock HJ, Jacob RA, Ames BN: Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc Natl Acad Sci U S A 1991;88:11003-11006. https://doi.org/10.1073/pnas.88.24.11003 |
||||
| 33 Jacob RA, Planalto FS, Agee RE: Cellular ascorbate depletion in healthy men. J Nutr 1992;122:1111-1118. https://doi.org/10.1093/jn/122.5.1111 |
||||
| 34 Kanďár R, Drábková P, Hampl R: The determination of ascorbic acid and uric acid in human seminal plasma using an HPLC with UV detection. J Chromatogr B Analyt Technol Biomed Life Sci 2011;879:2834-2839. https://doi.org/10.1016/j.jchromb.2011.08.007 |
||||
| 35 Thiele J, Freisleben HJ, Fuchs J, Ochsendorf F: Ascorbic acid and urate in human seminal plasma: Determination and interrelationships with chemiluminescence in washed semen. Hum Reprod 1995;10:110-115. https://doi.org/10.1093/humrep/10.1.110 |
||||
| 36 Abad C, Amengual MJ, Gosálvez J, Coward K, Hannaoui N, Benet J, García-Peiró A, Prats J: Effects of oral antioxidant treatment upon the dynamics of human sperm DNA fragmentation and subpopulations of sperm with highly degraded DNA. Andrologia 2013;45:211-216. https://doi.org/10.1111/and.12003 |
||||
| 37 Akmal M, Qadri JQ, Al-Waili N, Thangal S, Haq A, Saloom K: Improvement in Human Semen Quality After Oral Supplementation of Vitamin C. J Med Food 2006;9:440-442. https://doi.org/10.1089/jmf.2006.9.440 |
||||
| 38 Greco E, Iacobelli M, Rienzi L, Ubaldi F, Ferrero S, Tesarik J: Reduction of the incidence of sperm DNA fragmentation by oral antioxidant treatment. J Androl 2005;26:349-353. https://doi.org/10.2164/jandrol.04146 |
||||
| 39 Cyrus A, Kabir A, Goodarzi D, Moghimi M: The effect of adjuvant vitamin C after varicocele surgery on sperm quality and quantity in infertile men: a double blind placebo controlled clinical trial. Int Braz J Urol 2015;41:230-238. https://doi.org/10.1590/S1677-5538.IBJU.2015.02.07 |
||||
| 40 Mangoli E, Talebi AR, Anvari M, Taheri F, Vatanparast M, Rahiminia T, Hosseini A: Vitamin C attenuates negative effects of vitrification on sperm parameters, chromatin quality, apoptosis and acrosome reaction in neat and prepared normozoospermic samples. Taiwan J Obstet Gynecol 2018;57:200-204. https://doi.org/10.1016/j.tjog.2018.02.006 |
||||
| 41 Hughes CM, Lewis SE, McKelvey-Martin VJ, Thompson W: The effects of antioxidant supplementation during Percoll preparation on human sperm DNA integrity. Hum Reprod 1998;13:1240-1247. https://doi.org/10.1093/humrep/13.5.1240 |
||||
| 42 Fanaei H, Khayat S, Halvaei I, Ramezani V, Azizi Y, Kasaeian A, Mardaneh J, Parvizi MR, Akrami M: Effects of ascorbic acid on sperm motility, viability, acrosome reaction and DNA integrity in teratozoospermic samples. Iran J Reprod Med 2014;12:103-110. | ||||
| 43 Organization WH: WHO laboratory manual for the examination and processing of human semen. 2010 | ||||
| 44 Misro MM, Choudhury L, Upreti K, Gautam D, Chaki SP, Mahajan AS, Babbar R: Use of hydrogen peroxide to assess the sperm susceptibility to oxidative stress in subjects presenting a normal semen profile. Int J Androl 2004;27:82-87. https://doi.org/10.1046/j.0105-6263.2003.00451.x |
||||
| 45 Chaki SP, Misro MM: Assessment of human sperm function after hydrogen peroxide exposure. development of a vaginal contraceptive. Contraception 2002;66:187-192. https://doi.org/10.1016/S0010-7824(02)00349-9 |
||||
| 46 Rato L, Duarte AI, Tomás GD, Santos MS, Moreira PI, Socorro S, Cavaco JE, Alves MG, Oliveira PF: Pre-diabetes alters testicular PGC1-α/SIRT3 axis modulating mitochondrial bioenergetics and oxidative stress. Biochim Biophys Acta 2014;1837:335-344. https://doi.org/10.1016/j.bbabio.2013.12.008 |
||||
| 47 Benzie IF, Strain JJ: The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": the FRAP assay. Anal Biochem 1996;239:70-76. https://doi.org/10.1006/abio.1996.0292 |
||||
| 48 Moretti E, Mazzi L, Terzuoli G, Bonechi C, Iacoponi F, Martini S, Rossi C, Collodel G: Effect of quercetin, rutin, naringenin and epicatechin on lipid peroxidation induced in human sperm. Reprod Toxicol 2012;34:651-657. https://doi.org/10.1016/j.reprotox.2012.10.002 |
||||
| 49 Ben Abdallah F, Zribi N, Ammar‐Keskes L: Antioxidative potential of Quercetin against hydrogen peroxide induced oxidative stress in spermatozoa in vitro . Andrologia 2011;43:261-265. https://doi.org/10.1111/j.1439-0272.2010.01063.x |
||||
| 50 Benzie IFF, Devaki M: The ferric reducing/antioxidant power (FRAP) assay for non-enzymatic antioxidant capacity: concepts, procedures, limitations and applications; in Apak R, Çapanoglu E, Shahidi F (eds): Measurement of Antioxidant Activity & Capacity, Wiley, 2018, pp 77-106. https://doi.org/10.1002/9781119135388.ch5 |
||||
| 51 Ahmad G, Agarwal A, Esteves SC, Sharma R, Almasry M, Al-Gonaim A, AlHayaza G, Singh N, Al Kattan L, Sannaa WM, Sabanegh E: Ascorbic acid reduces redox potential in human spermatozoa subjected to heat-induced oxidative stress. Andrologia 2017;49:e12773. https://doi.org/10.1111/and.12773 |
||||
| 52 Karabulut S, Korkmaz O, Erdem Altun C, Demiroğlu Zergeroǧlu A, Keskin İ: Quercetin enhances human sperm motility in a dose and time dependent manner. Acta Pharm Sci 2020; DOI: 10.23893/1307-2080.APS.05810. https://doi.org/10.23893/1307-2080.APS.05810 |
||||
| 53 Evdokimov VV, Barinova KV, Turovetskii VB, Muronetz VI, Schmalhausen EV: Low concentrations of hydrogen peroxide activate the antioxidant defense system in human sperm cells. Biochemistry (Moscow) 2015;80:1178-1185. https://doi.org/10.1134/S0006297915090084 |
||||
| 54 Kurkowska W, Bogacz A, Janiszewska M, Gabryś E, Tiszler M, Bellanti F, Kasperczyk S, Machoń-Grecka A, Dobrakowski M, Kasperczyk A: Oxidative Stress is Associated with Reduced Sperm Motility in Normal Semen. Am J Mens Health 2020;14:1557988320939731. https://doi.org/10.1177/1557988320939731 |
||||
| 55 Walters JLH, De Iuliis GN, Nixon B, Bromfield EG: Oxidative Stress in the Male Germline: A Review of Novel Strategies to Reduce 4-Hydroxynonenal Production. Antioxidants (Basel) 2018;7:132. https://doi.org/10.3390/antiox7100132 |
||||
| 56 Moazamian R, Polhemus A, Connaughton H, Fraser B, Whiting S, Gharagozloo P, Aitken R: Oxidative stress and human spermatozoa: Diagnostic and functional significance of aldehydes generated as a result of lipid peroxidation. Mol Hum Reprod 2015;21:502-515. https://doi.org/10.1093/molehr/gav014 |
||||
| 57 Nowicka-Bauer K, Nixon: Molecular Changes Induced by Oxidative Stress that Impair Human Sperm Motility. Antioxidants 2020;9:134. https://doi.org/10.3390/antiox9020134 |
||||
| 58 Raad G, Mansour J, Ibrahim R, Azoury J, Azoury J, Mourad Y, Fakih C, Azoury J: What are the effects of vitamin C on sperm functional properties during direct swim-up procedure? Zygote 2019;27:69-77. https://doi.org/10.1017/S0967199419000030 |
||||
| 59 Li Z, Lin Q, Liu R, Xiao W, Liu W: Protective effects of ascorbate and catalase on human spermatozoa during cryopreservation. J Androl 2010;31:437-444. https://doi.org/10.2164/jandrol.109.007849 |
||||
| 60 du Plessis SS, Agarwal A, Mohanty G, van der Linde M: Oxidative phosphorylation versus glycolysis: what fuel do spermatozoa use? Asian J Androl 2015;17:230-235. https://doi.org/10.4103/1008-682X.135123 |
||||
| 61 Nascimento JM, Shi LZ, Tam J, Chandsawangbhuwana C, Durrant B, Botvinick EL, Berns MW: Comparison of glycolysis and oxidative phosphorylation as energy sources for mammalian sperm motility, using the combination of fluorescence imaging, laser tweezers, and real-time automated tracking and trapping. J Cell Physiol 2008;217:745-751. https://doi.org/10.1002/jcp.21549 |
||||
| 62 Gallon F, Marchetti C, Jouy N, Marchetti P: The functionality of mitochondria differentiates human spermatozoa with high and low fertilizing capability. Fertil Steril 2006;86:1526-1530. https://doi.org/10.1016/j.fertnstert.2006.03.055 |
||||
| 63 Ferramosca A, Provenzano SP, Coppola L, Zara V: Mitochondrial Respiratory Efficiency is Positively Correlated With Human Sperm Motility. Urology 2012;79:809-814. https://doi.org/10.1016/j.urology.2011.12.042 |
||||
| 64 Gray LR, Tompkins SC, Taylor EB: Regulation of pyruvate metabolism and human disease. Cell Mol Life Sci 2014;71:2577-2604. https://doi.org/10.1007/s00018-013-1539-2 |
||||
| 65 Mitchell M, Cashman KS, Gardner DK, Thompson JG, Lane M: Disruption of Mitochondrial Malate-Aspartate Shuttle Activity in Mouse Blastocysts Impairs Viability and Fetal Growth1. Biol Reprod 2009;80:295-301. https://doi.org/10.1095/biolreprod.108.069864 |
||||
| 66 Newsholme P, Keane K, Homem de Bittencourt Jr P, Krause M: The Impact of Inflammation on Pancreatic β-Cell Metabolism, Function and Failure in T1DM and T2DM: Commonalities and Differences, in Escher AP, Li A (eds): Type 1 Diabetes, InTech, 2013, pp 127-165. https://doi.org/10.5772/55349 |
||||
| 67 Deburgos NG, Gallina F, Burgos C, Blanco A: Effect of L-malate on pyruvate-dehydrogenase activity of spermatozoa. Arch Biochem Biophys 1994;308:520-524. https://doi.org/10.1006/abbi.1994.1073 |
||||
| 68 Storey BT, Kayne FJ: Energy metabolism of spermatozoa. VII. Interactions between lactate, pyruvate and malate as oxidative substrates for rabbit sperm mitochondria. Biol Reprod 1978;18:527-536. https://doi.org/10.1095/biolreprod18.4.527 |
||||
| 69 Bose S, Ramesh V, Locasale JW: Acetate Metabolism in Physiology, Cancer, and Beyond. Trends Cell Biol 2019;29:695-703. https://doi.org/10.1016/j.tcb.2019.05.005 |
||||
| 70 Alves M, Socorro S, Silva J, Barros A, Sousa M, Cavaco J, Oliveira PJBeba: In vitro cultured human Sertoli cells secrete high amounts of acetate that is stimulated by 1:²-estradiol and suppressed by insulin deprivation. Biochim Biophys Acta 2012;1823 8:1389-1394. https://doi.org/10.1016/j.bbamcr.2012.06.002 |
||||
| 71 Carrageta DF, Guerra-Carvalho B, Sousa M, Barros A, Oliveira PF, Monteiro MP, Alves MG: Mitochondrial Activation and Reactive Oxygen-Species Overproduction during Sperm Capacitation are Independent of Glucose Stimuli. Antioxidants 2020;9 https://doi.org/10.3390/antiox9080750 |
||||
| 72 Zribi N, Chakroun NF, Ben Abdallah F, Elleuch H, Sellami A, Gargouri J, Rebai T, Fakhfakh F, Keskes LA: Effect of freezing-thawing process and quercetin on human sperm survival and DNA integrity. Cryobiology 2012;65:326-331. https://doi.org/10.1016/j.cryobiol.2012.09.003 |
||||
| 73 Seifi-Jamadi A, Kohram H, Zare Shahneh A, Ansari M, Macías-García B: Quercetin Ameliorate Motility in Frozen-Thawed Turkmen Stallions Sperm. J Equine Vet Sci 2016;45:73-77. https://doi.org/10.1016/j.jevs.2016.06.078 |
||||
| 74 Najafi A, Kia HD, Mehdipour M, Hamishehkar H, Álvarez-Rodríguez M: Effect of quercetin loaded liposomes or nanostructured lipid carrier (NLC) on post-thawed sperm quality and fertility of rooster sperm. Theriogenology 2020;152:122-128. https://doi.org/10.1016/j.theriogenology.2020.04.033 |
||||
| 75 Avdatek F, Yeni D, İnanç ME, Çil B, Tuncer BP, Türkmen R, Taşdemir U: Supplementation of quercetin for advanced DNA integrity in bull semen cryopreservation. Andrologia 2018;50:e12975. https://doi.org/10.1111/and.12975 |
||||
| 76 Sierens J, Hartley J, Campbell M, Leathem A, Woodside J: In vitro isoflavone supplementation reduces hydrogen peroxide‐induced DNA damage in sperm. Teratog Carcinog Mutagen 2002;22:227-234. https://doi.org/10.1002/tcm.10015 |
||||
| 77 Calvert SJ, Reynolds S, Paley MN, Walters SJ, Pacey AA: Probing human sperm metabolism using 13C-magnetic resonance spectroscopy. Mol Hum Reprod 2019;25:30-41. https://doi.org/10.1093/molehr/gay046 |
||||
| 78 Reynolds S, Ismail NFb, Calvert SJ, Pacey AA, Paley MNJ: Evidence for Rapid Oxidative Phosphorylation and Lactate Fermentation in Motile Human Sperm by Hyperpolarized 13C Magnetic Resonance Spectroscopy. Sci Rep 2017;7:4322. https://doi.org/10.1038/s41598-017-04146-1 |
||||
| 79 Guarino VA, Oldham WM, Loscalzo J, Zhang Y-Y: Reaction rate of pyruvate and hydrogen peroxide: assessing antioxidant capacity of pyruvate under biological conditions. Sci Rep 2019;9:19568. https://doi.org/10.1038/s41598-019-55951-9 |
||||
| 80 Desagher S, Glowinski J, Prémont J: Pyruvate protects neurons against hydrogen peroxide-induced toxicity. J Neurosci 1997;17:9060-9067. https://doi.org/10.1523/JNEUROSCI.17-23-09060.1997 |
||||
| 81 Babich H, Liebling E, Burger R, Zuckerbraun H, Schuck A: Choice of DMEM, formulated with or without pyruvate, plays an important role in assessing the in vitro cytotoxicity of oxidants and prooxidant nutraceuticals. In vitro Cell Dev Biol Anim 2009;45:226-233. https://doi.org/10.1007/s11626-008-9168-z |
||||
| 82 Sheraz M, Khan M, Ahmed S, Kazi S, Ahmad I: Stability and Stabilization of Ascorbic Acid. Househ Pers Care Today 2015;10:20-25. | ||||
| 83 Gallarate M, Carlotti ME, Trotta M, Bovo S: On the stability of ascorbic acid in emulsified systems for topical and cosmetic use. Int J Pharm 1999;188:233-241. https://doi.org/10.1016/S0378-5173(99)00228-8 |
||||
| 84 Dolińska B, Ostróżka-Cieślik A, Caban A, Rimantas K, Leszczyńska L, Ryszka F: Influence of trace elements on stabilization of aqueous solutions of ascorbic acid. Biol Trace Elem Res 2012;150:509-512. https://doi.org/10.1007/s12011-012-9524-4 |
||||
| 85 Shi S, Long R, Tang C, Yang z, Fu Q, Xu J, Tong C, Guo Y, Wang D: A natural hyperoside based novel light-up fluorescent probe with AIE and ESIPT characteristics for on-site and long-term imaging of β-galactosidase in living cells. J Mater Chem C 2020;8:11860-11865. https://doi.org/10.1039/D0TC01981J |
||||