×
![]()
Corresponding Author: Bonnie Blazer-Yost
Biology Department, Indiana University Purdue University Indianapolis, 723 West Michigan Street, Indianapolis, IN 46228 (USA)
Tel. +1-317-278-1145, Fax +1-317-274-2846, E-Mail bblazer@iupui.edu
Channels and Transporters in Astrocyte Volume Regulation in Health and Disease
Makenna M. Reed Bonnie Blazer-Yost
Indiana University Purdue University Indianapolis, Indianapolis, IN, USA
Introduction
The extracellular fluids of the central nervous system (CNS) are comprised of blood, interstitial fluid (ISF) and cerebrospinal fluid (CSF). Collectively, these are responsible for functions such as hormone and cytokine signaling, CNS environment homeostasis, nutrient delivery, brain waste clearance, and brain cushioning. The composition of each of the fluids is distinct and the movement of water, small molecular weight components and even cells between compartments represent highly controlled processes [1-4]. There are several cell types that regulate the composition, movement, and clearance of the three different fluids. The choroid plexus secretes CSF into the brain’s ventricular system where it is absorbed, dispersed, or drained by other cell types such as ependymal, meningeal, lymphatic, and glial cells [5]. The blood-brain barrier is one of the least permeable vascular systems in the body, although specific transport across this barrier does take place and disruption of the barrier is a common pathological occurrence in disease or injury [6-10]. On a more localized level, maintenance of the fluid composition, particularly in niches such as the perisynaptic domain and the perivascular space, is controlled by specialized cells called astrocytes [11].
Astrocytes are one of the most common CNS glial cells and are found throughout the brain. Astrocyte function depends on specific localization within the brain. Of importance are astrocytes found at the blood brain barrier (BBB), where they play a key role in maintenance of CSF and ISF [11]. Astrocytes form a layer adjacent to the pia mater called the glia limitans. The main function of these astrocytes is protection and regulation of CSF adjacent to pial vessels [12, 13]. Alternatively, astrocytes with endfeet in close proximity to synaptic junctions, support neurons through maintenance of a constant extracellular milieu during neuronal activity, playing a supporting role in synaptic transmission and modulating information via the secretion of gliotransmitters [14-19]. Perivascular astrocytes are important in glymphatic function. The glymphatic system was first proposed as method of cerebrospinal fluid (CSF) and interstitial fluid (ISF) exchange and clearance [20]. CSF enters the periarterial space where it can be taken up by astrocytes and transported throughout the brain parenchyma to perivenous spaces and then drained through cervical lymphatics [20-24]. The glymphatic system has been proposed to be important in the clearance of brain waste and toxic materials, including amyloid β [20].
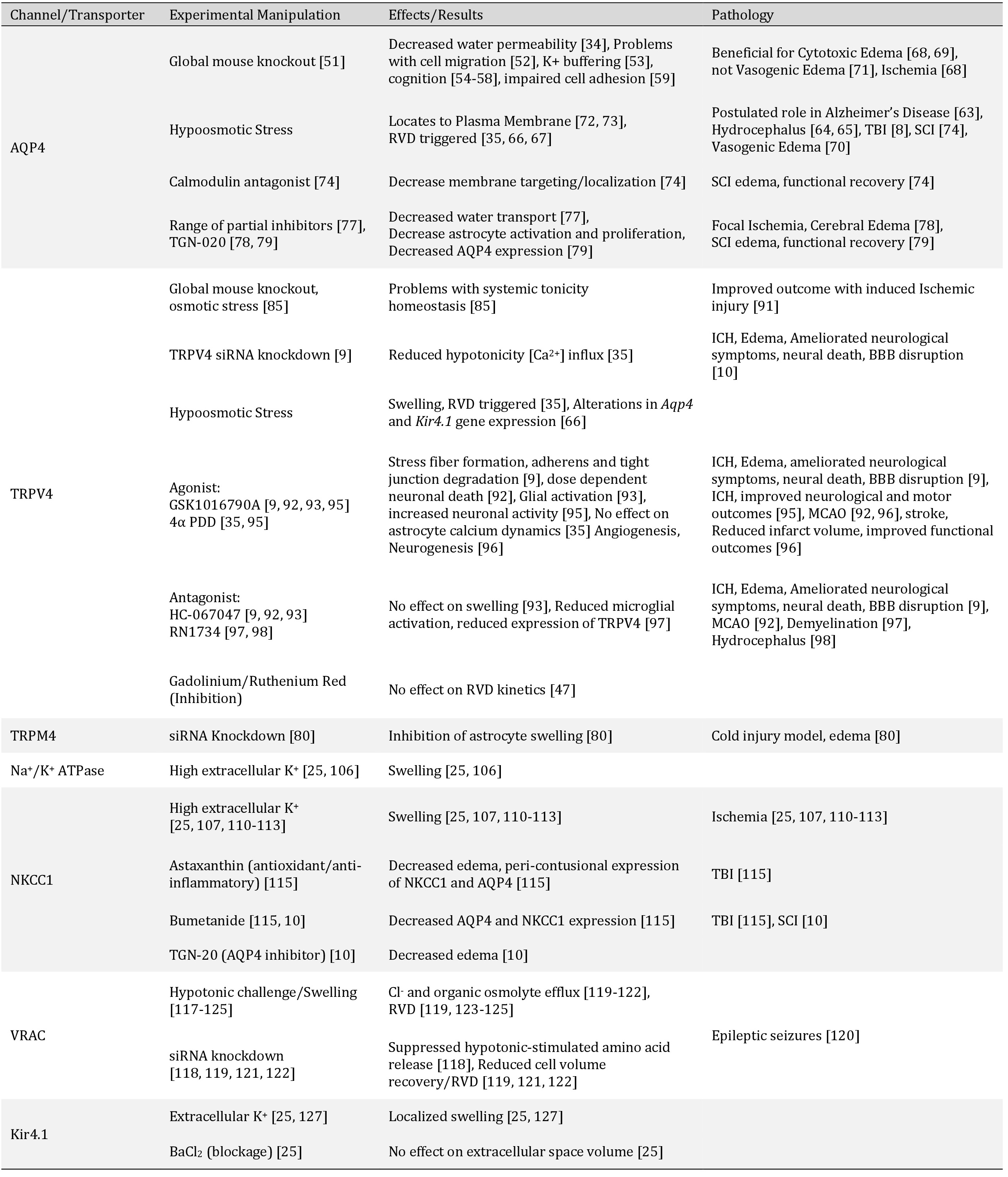
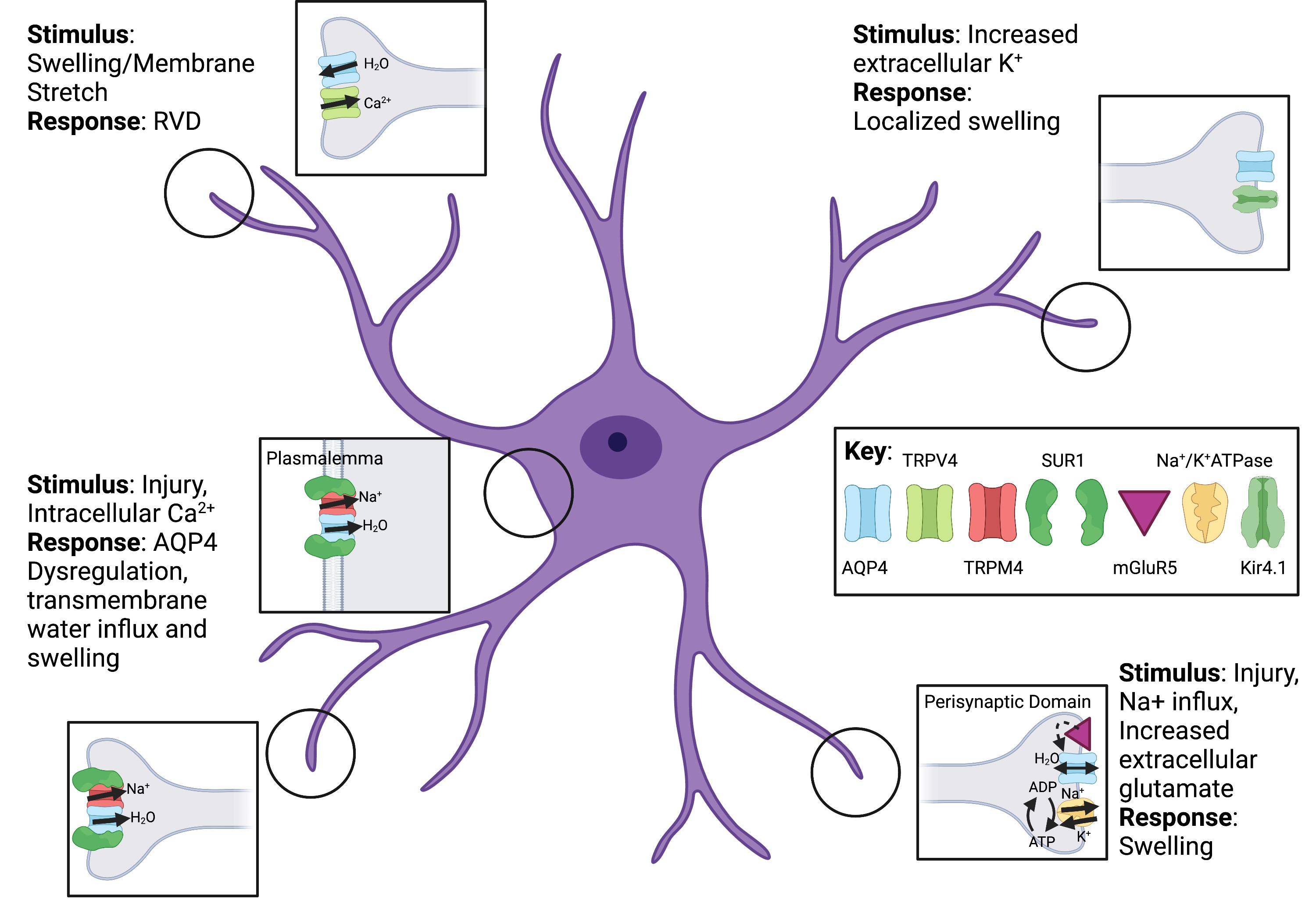
In this review, we will focus on the transporters involved in the maintenance of the extracellular milieu and, as a consequence, cell volume in astrocytes under normal and pathological conditions (Table 1). Astrocytes are responsible for maintaining normal brain environment including electrolyte and fluid balance, which involves the uptake and removal of extracellular compounds. Notable in this regard is the removal of accumulating extracellular K+ during times of high neuronal activity [25]. Glial cells in general, and astrocytes in particular, swell in response to a variety of injuries including ischemic injury, stroke, traumatic brain injury and inflammation [6-7]. Excitotoxicity, neuronal and vascular degeneration, and metabolic disruption can all be a result of detrimental glial swelling [26, 27]. Depending on severity, these processes can either be followed by a normal decrease in volume or abnormal function and cell death [28, 29].
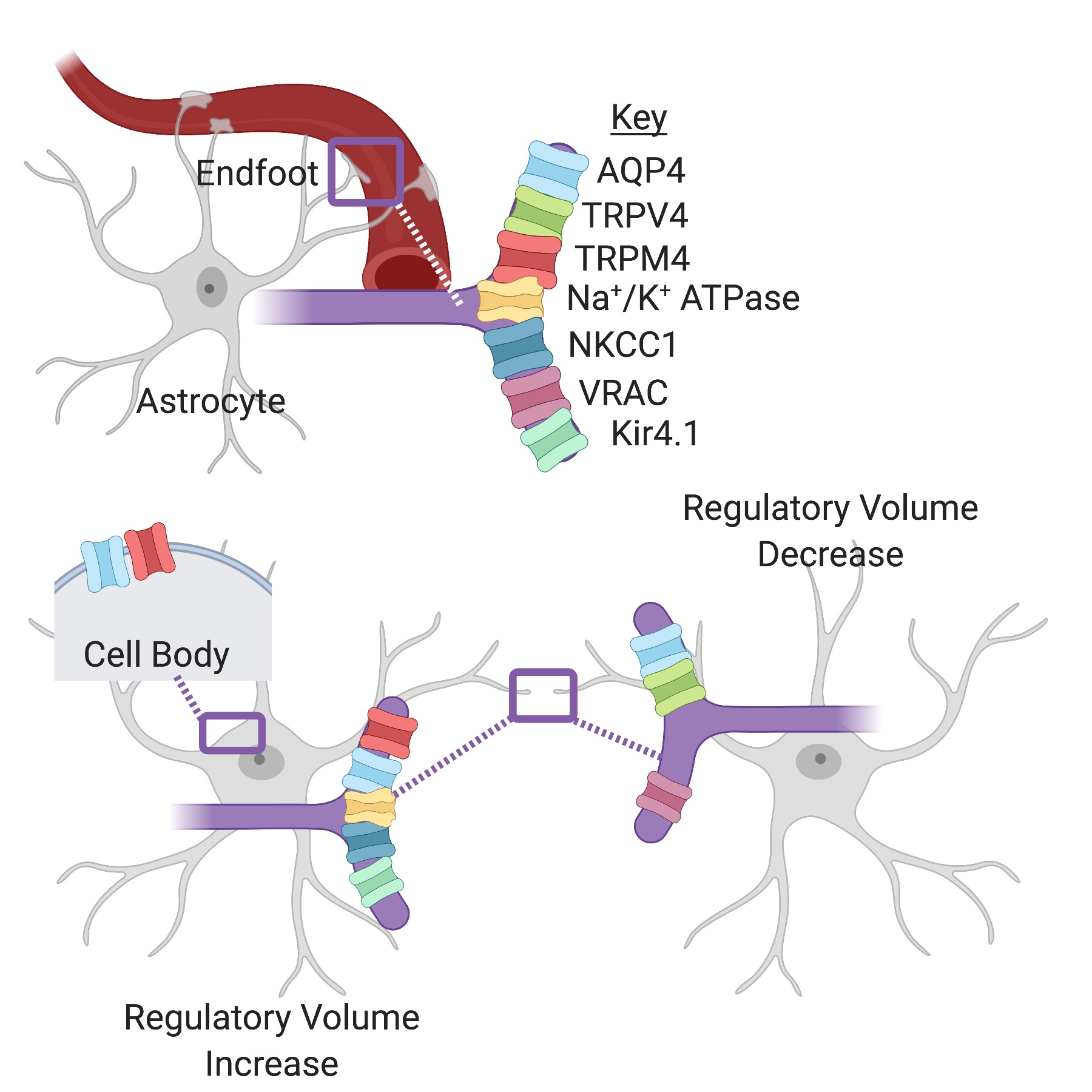
Regulatory cell volume changes are mediated by transmembrane ion fluxes [30, 31]. Astrocytes contain key channels and transporters that are involved in either regulatory volume increase (RVI) or decrease (RVD) (Fig. 1). During RVD ions efflux from the cell, while during RVI, ions accumulate within the cell to maintain osmotic equilibrium. Simultaneous activation of these mechanisms is avoided through inhibition of the opposing mechanism [30, 32, 33]. The astrocytic channels and transporters that are currently implicated in cell volume regulation include the bidirectional water transporter, aquaporin-4 (AQP4), the osmo- and mechano-sensitive channel, transient receptor potential cation channel, subfamily vanilloid, member 4 (TRPV4), the sulfonylurea receptor 1 (SUR1)-transient receptor potential melastatin 4 (TRPM4) complex, the volume-regulated anion channel (VRAC), the inwardly rectifying potassium channel (Kir4.1), sodium potassium adenosine triphosphatase pump (Na+/K+ ATPase) and Na+-K+-Cl- cotransporter (NKCC1) (Fig. 2).
The authors would like to thank our colleagues, Casandra Carrillo, Alexandra Hochstetler, and Louise Hulme for discussions and comments on the manuscript.
Author Contributions
Both authors contributed to the writing and editing of the manuscript.
Funding
Research in the authors’ laboratory was funded by a Hydrocephalus Association/ Team Hydro Innovator Award; The Office of the Assistant Secretary of Defense for Health Affairs, through the Peer Reviewed Medical Research Program, Investigator Initiated Research Award No. W81XWH-17-1-0537; and a Mayfield Education and Research Foundation Award.
Statement of Ethics
The authors have no ethical conflicts to disclose.
The authors have no conflicts of interest to declare.
| 1 Millen JW, Woollam DHM: The Anatomy of the Cerebrospinal Fluid. London, Oxford University Press, 1962. | ||||
| 2 Davson H, Segal MB: Physiology of the CSF and Blood-brain Barriers. Boca Raton/London, CRC Press, 1996. | ||||
| 3 Bito LZ, Davson H: Local variations in cerebrospinal fluid composition and its relationship to the composition of the extracellular fluid of the cortex. Exp Neurol 1966;14:264-280. https://doi.org/10.1016/0014-4886(66)90114-2 |
||||
| 4 Merritt HH, Fremont-Smith F: The Cerebrospinal Fluid. Philadelphia/London, WB Saunders, 1937. | ||||
| 5 Hladky SB, Barrand MA: Mechanisms of fluid movement into, through and out of the brain: evaluation of the evidence. Fluids Barriers CNS 2014;11:26. https://doi.org/10.1186/2045-8118-11-26 |
||||
| 6 Dostovic Z, Dostovic E, Smajlovic D, Ibrahimagic OC, Avdic L: Brain Edema After Ischaemic Stroke. Med Arch 2016;70:339-341. https://doi.org/10.5455/medarh.2016.70.339-341 |
||||
| 7 Unterberg AW, Stover J, Kress B, Kiening KL: Edema and brain trauma. Neurosci 2004;129:1021-1029. https://doi.org/10.1016/j.neuroscience.2004.06.046 |
||||
| 8 Neri M, Frati A, Turillazzi E, Cantatore S, Cipolloni L, Di Paolo M, Frati P, La Russa R, Maiese A, Scopetti M, Santurro A, Sessa F, Zamparese R, Fineschi V: Immunohistochemical Evaluation of Aquaporin-4 and its Correlation with CD68, IBA-1, HIF-1α, GFAP, and CD15 Expressions in Fatal Traumatic Brain Injury. Int J Mol Sci 2018;19:3544. https://doi.org/10.3390/ijms19113544 |
||||
| 9 Zhao H, Zhang K, Tang R, Meng H, Zou Y, Wu P, Hu R, Liu X, Feng H, Chen Y: TRPV4 blockade preserves the blood-brain barrier by inhibiting stress fiber formation in a rat model of intracerebral hemorrhage. Front Mol Neurosci 2018;11:97. https://doi.org/10.3389/fnmol.2018.00097 |
||||
| 10 Yan X, Liu J, Wang X, Li W, Chen J, Sun H: Pretreatment with AQP4 and NKCC1 Inhibitors Concurrently Attenuated Spinal Cord Edema and Tissue Damage after Spinal Cord Injury in Rats. Front Physiol 2018;9:6. https://doi.org/10.3389/fphys.2018.00006 |
||||
| 11 Simard M, Nedergaard M: The neurobiology of glia in the context of water and ion homeostasis. Neurosci 2004;129:877-896. https://doi.org/10.1016/j.neuroscience.2004.09.053 |
||||
| 12 Alvarez JI, Dodelet-Devillers A, Kebir H, Ifergan I, Fabre PJ, Terouz S, Sabbagh M, Wosik K, Bourbonniere L, Bernard M, van Horssen J, de Vries HE, Charron F, Prat A: The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence. Science 2011;334:1727-1731. https://doi.org/10.1126/science.1206936 |
||||
| 13 Obermeier B, Daneman R, Ransohoff RM: Development, maintenance and disruption of the blood-brain barrier. Nat Med 2013;19:1584-1596. https://doi.org/10.1038/nm.3407 |
||||
| 14 Allen NJ, Barres BA: Neuroscience: Glia - more than just brain glue. Nature 2009;457:675-677. https://doi.org/10.1038/457675a |
||||
| 15 Allen NJ, Bennett ML, Foo LC, Wang GX, Chakraborty C, Smith SJ, Barres BA: Astrocyte glypicans 4 and 6 promote formation of excitatory synapses via GluA1 AMPA receptors. Nature 2012;486:410-414. https://doi.org/10.1038/nature11059 |
||||
| 16 Chung WS, Clarke LE, Wang GX, Stafford BK, Sher A, Chakraborty C, Joung J, Foo LC, Thompson A, Chen C, Smith SJ, Barres BA: Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature 2013;504:394-400. https://doi.org/10.1038/nature12776 |
||||
| 17 Khakh BS, Sofroniew MV: Diversity of astrocyte functions and phenotypes in neural circuits. Nat Neurosci 2015;18:942-952. https://doi.org/10.1038/nn.4043 |
||||
| 18 Molofsky AV, Kelley KW, Tsai H, Redmond SA, Chang SM, Madireddy L, Chan JR, Baranzini SE, Ullian EM, Rowitch DH: Astrocyte-encoded positional cues maintain sensorimotor circuit integrity. Nature 2014;509:189-194. https://doi.org/10.1038/nature13161 |
||||
| 19 Tsai HH, Li H, Fuentealba LC, Molofsky AV, Taveira-Marques R, Zhuang H, Tenney A, Murnen AT, Fancy SPJ, Merkle F, Kessaris N, Alvarez-Buylla A, Richardson WD, Rowitch DH: Regional astrocyte allocation regulates CNS synaptogenesis and repair. Science 2012;337:358-362. https://doi.org/10.1126/science.1222381 |
||||
| 20 Iliff JJ, Wang M, Liao Y, Plogg BA, Peng W, Gundersen GA, Benveniste H, Vates GE, Deane R, Goldman SA, Nagelhus EA, Nedergaard M: A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci Transl Med 2012;4:147ra111. https://doi.org/10.1126/scitranslmed.3003748 |
||||
| 21 Jessen NA, Munk AS, Lundgaard I, Nedergaard M: The Glymphatic System: A Beginner's Guide. Neurochem Res 2015;40:2583-2599. https://doi.org/10.1007/s11064-015-1581-6 |
||||
| 22 Iliff JJ, Nedergaard M: Is there a cerebral lymphatic system?. Stroke 2013;44:S93-S95. https://doi.org/10.1161/STROKEAHA.112.678698 |
||||
| 23 Johnston M, Zakharov A, Papaiconomou C, Salmasi G, Armstrong D: Evidence of connections between cerebrospinal fluid and nasal lymphatic vessels in humans, non-human primates and other mammalian species. Cerebrospinal Fluid Res 2004;1:2. https://doi.org/10.1186/1743-8454-1-2 |
||||
| 24 Murtha LA, Yang Q, Parsons MW, Levi CR, Beard DJ, Spratt NJ, McLeod DD: Cerebrospinal fluid is drained primarily via the spinal canal and olfactory route in young and aged spontaneously hypertensive rats. Fluids Barriers CNS 2014;11:12. https://doi.org/10.1186/2045-8118-11-12 |
||||
| 25 Larsen BR, Assentoft M, Cotrina ML, Hua SZ, Nedergaard M, Kaila K, Voipio J, MacAulay N: Contributions of the Na⁺/K⁺-ATPase, NKCC1, and Kir4.1 to hippocampal K⁺ clearance and volume responses. Glia 2014;62:608-622. https://doi.org/10.1002/glia.22629 |
||||
| 26 Häussinger D, Kircheis G, Fischer R, Schliess F, vom Dahl S: Hepatic encephalopathy in chronic liver disease: a clinical manifestation of astrocyte swelling and low-grade cerebral edema?. J Hepatol 2000;32:1035-1038. https://doi.org/10.1016/S0168-8278(00)80110-5 |
||||
| 27 Hirrlinger PG, Wurm A, Hirrlinger J, Bringmann A, Reichenbach A: Osmotic swelling characteristics of glial cells in the murine hippocampus, cerebellum, and retina in situ. J Neurochem 2008;105:1405-1417. https://doi.org/10.1111/j.1471-4159.2008.05243.x |
||||
| 28 Kempski O: Cerebral edema. Semin Nephrol 2001;21:303-307. https://doi.org/10.1053/snep.2001.21665 |
||||
| 29 Stokum JA, Kurland DB, Gerzanich V, Simard JM: Mechanisms of Astrocyte-Mediated Cerebral Edema. Neurochem Res 2015;40:317-328. https://doi.org/10.1007/s11064-014-1374-3 |
||||
| 30 Lang F, Busch GL, Ritter M, Völkl H, Waldegger S, Gulbins E, Häussinger Dz: Functional significance of cell volume regulatory mechanisms. Physiol Rev 1998;78:247-306. https://doi.org/10.1152/physrev.1998.78.1.247 |
||||
| 31 Verbalis JG, SR Gullans: Hyponatremia causes large sustained reductions in brain content of multiple organic osmolytes in rats. Brain Res 1991;567:274-282. https://doi.org/10.1016/0006-8993(91)90806-7 |
||||
| 32 Palfrey HC: Protein phosphorylation control in the activity of volume-sensitive transport systems; in Kevin Strange (eds): Cellular and Molecular Physiology of Cell Volume Regulation. Boca Raton/London, CRC Press, 1994, pp 201-214. https://doi.org/10.1201/9780367812140-15 |
||||
| 33 Parker JC: Coordinated regulation of volume-activated transport pathways; in Kevin Strange (eds): Cellular and Molecular Physiology of Cell Volume Regulation. Boca Raton/London, CRC Press, 1994, pp 311-324. https://doi.org/10.1201/9780367812140-21 |
||||
| 34 Solenov E, Watanabe H, Manley GT, Verkman AS: Sevenfold-reduced osmotic water permeability in primary astrocyte cultures from AQP-4-deficient mice, measured by a fluorescence quenching method. Am J Physiol Cell Physiol 2004;286:C426-C432. https://doi.org/10.1152/ajpcell.00298.2003 |
||||
| 35 Benfenati V, Caprini M, Dovizio M, Mylonakou MN, Ferroni S, Petter Ottersen O, Amiry-Moghaddam M: An aquaporin-4/transient receptor potential vanilloid 4 (AQP4/TRPV4) complex is essential for cell-volume control in astrocytes. Proc Natl Acad Sci U S A 2011;108:2563-2568. https://doi.org/10.1073/pnas.1012867108 |
||||
| 36 Nielsen S, Nagelhus EA, Amiry-Moghaddam M, Bourque C, Agre P, Otterson OP: Specialized membrane domains for water transport in glial cells: High-resolution immunogold cytochemistry of aquaporin-4 in rat brain. J Neurosci 1997;17:171-180. https://doi.org/10.1523/JNEUROSCI.17-01-00171.1997 |
||||
| 37 Rash JE, Yasumura T, Hudson CS, Agre P, Nielsen S: Direct immunogold labeling of aquaporin-4 in square arrays of astrocyte and ependymocyte plasma membranes in rat brain and spinal cord. Proc Natl Acad Sci
U S A 1998;95:11981-11986. https://doi.org/10.1073/pnas.95.20.11981 |
||||
| 38 Amiry-Moghaddam M, Petter Ottersen O: The molecular basis of water transport in the brain. Nat Rev Neurosci 2003;4:991-1001. https://doi.org/10.1038/nrn1252 |
||||
| 39 Verbalis JG: Control of brain volume during hypoosmolality and hyperosmolality. Adv Exp Med Biol 2006; DOI: 10.1007/0-387-30172-0_8. https://doi.org/10.1007/0-387-30172-0_8 |
||||
| 40 Amiry-Moghaddam M, Frydenlund DS, Petter Ottersen O: Anchoring of aquaporin-4 in brain: Molecular mechanisms and implications for the physiology and pathophysiology of water transport. Neurosci 2004;129:999-1010. https://doi.org/10.1016/j.neuroscience.2004.08.049 |
||||
| 41 Moe SE, Sorbo JG, Sogaard R, Zeuthen T, Petter Ottersen O, Holen T: New isoforms of rat Aquaporin-4. Genom 2008;91:367-377. https://doi.org/10.1016/j.ygeno.2007.12.003 |
||||
| 42 Jung JS, Bhat RV, Preston GM, Guggino WB, Baraban JM, Agre P: Molecular characterization of an aquaporin cDNA from brain: Candidate osmoreceptor and regulator of water balance. Proc Natl Acad Sci U S A 1994;91:13052-13056. https://doi.org/10.1073/pnas.91.26.13052 |
||||
| 43 Hasegawa H, Ma T, Skach W, Matthay MA, Verkman AS: Molecular cloning of a mercurial-insensitive water channel expressed in selected water-transporting tissues. J Biol Chem 1994;269:5497-5500. https://doi.org/10.1016/S0021-9258(17)37486-0 |
||||
| 44 Neely JD, Christensen BM, Nielsen S, Agre P: Heterotetrameric composition of aquaporin-4 water channels. Biochem 1999;38:11156-11163. https://doi.org/10.1021/bi990941s |
||||
| 45 Lu M, Lee MD, Smith BL, Jung JS, Agre P, Verdijk MA, Merkx G, Rijss JP, Deen PM: The human AQP4 gene: Definition of the locus encoding two water channel polypeptides in brain. Proc Natl Acad Sci U S A 1996;93:10908-10912. https://doi.org/10.1073/pnas.93.20.10908 |
||||
| 46 Yang B, Ma T, Verkman AS: cDNA cloning, gene organization, and chromosomal localization of a human mercurial insensitive water channel: Evidence for distinct transcriptional units. J Biol Chem 1995;270:22907-22913. https://doi.org/10.1074/jbc.270.39.22907 |
||||
| 47 Mola M, Sparaneo A, Gargano C, Spray DC, Svelto M, Frigeri A, Scemes E, Nicchia G: The speed of swelling kinetics modulates cell volume regulation and calcium signaling in astrocytes: A different point of view on the role of aquaporins. Glia 2016;64:139-154. https://doi.org/10.1002/glia.22921 |
||||
| 48 Potokar M, Stenovec M, Jorgacevski J, Holen T, Kreft M, Otterson OP, Zorec R: Regulation of AQP4 surface expression via vesicle mobility in astrocytes. Glia 2013;61:917-928. https://doi.org/10.1002/glia.22485 |
||||
| 49 Lisjak M, Potokar M, Rituper B, Jorgačevski J, Zorec R: AQP4e-Based Orthogonal Arrays Regulate Rapid Cell Volume Changes in Astrocytes. J Neurosci 2017;37:10748-10756. https://doi.org/10.1523/JNEUROSCI.0776-17.2017 |
||||
| 50 Lisjak M, Potokar M, Zorec R, Jorgačevski J: Indirect Role of AQP4b and AQP4d Isoforms in Dynamics of Astrocyte Volume and Orthogonal Arrays of Particles. Cells 2020;9:735. https://doi.org/10.3390/cells9030735 |
||||
| 51 Ma T, Yang B, Gillespie A, Carlson EJ, Epstein CJ, Verkman AS: Generation and phenotype of a transgenic knockout mouse lacking the mercurial-insensitive water channel aquaporin-4. J Clin Invest 1997;100:957-962. https://doi.org/10.1172/JCI231 |
||||
| 52 Saadoun S, Papadopoulos MC, Watanabe H, Yan D, Manley GT, Verkman AS: Involvement of aquaporin-4 in astroglial cell migration and glial scar formation. J Cell Sci 2005;118:5691-5698. https://doi.org/10.1242/jcs.02680 |
||||
| 53 Binder DK, Yao X, Verkman AS, Manley GT: Increased seizure duration in mice lacking aquaporin-4 water channels. Acta Neurochir Suppl 2006;96:389-392. https://doi.org/10.1007/3-211-30714-1_80 |
||||
| 54 Skucas VA, Mathews IB, Yang J, Cheng Q, Treister A, Duffy AM, Scharfman HE: Impairment of select forms of spatial memory and neurtrophin-dependent synaptic plasticity by deletion of glial aquaporin-4. J Neurosci 2011;31:6392-6397. https://doi.org/10.1523/JNEUROSCI.6249-10.2011 |
||||
| 55 Scharfman HE, Binder DK: Aquaporin-4 water channels and synaptic plasticity in the hippocampus. Neurochem Int 2013;63:702-711. https://doi.org/10.1016/j.neuint.2013.05.003 |
||||
| 56 Fan Y, Zhang J, Sun XL, Gao L, Zeng XN, Ding JH, Cao C, Niu L, Hu G: Sex- and region-specific alterations of basal amino acid and monoamine metabolism in the brain of aquaporin-4 knockout mice. J Neurosci Res 2005;82:458-464. https://doi.org/10.1002/jnr.20664 |
||||
| 57 Zeng XN, Sun XL, Gao L, Fan Y, Ding JH, Hu G: Aquaporin-4 deficiency down-regulates glutamate uptake and GLT-1 expression in astrocytes. Mol Cell Neurosci 2007;34:34-39. https://doi.org/10.1016/j.mcn.2006.09.008 |
||||
| 58 Li YK, Wang F, Wang W, Luo Y, Wu PF, Xiao JL, Hu ZL, Jin Y, Hu G, Chen JG: Aquaporin-4 deficiency impairs synaptic plasticity and associative fear memory in the lateral amygdala: involvement of downregulation of glutamate transporter-1 expression. Neuropsychopharmacol 2012;37:1867-1878. https://doi.org/10.1038/npp.2012.34 |
||||
| 59 Hiroaki Y, Tani K, Kamegawa A, Gyobu N, Nishikawa K, Suzuki H, Walz T, Sasaki S, Mitsuoka K, Kimura K, Mizoguchi A, Fujiyoshi Y: Implications of the aquaporin-4 structure on array formation and cell adhesion.
J Mol Biol 2006;355:628-639. https://doi.org/10.1016/j.jmb.2005.10.081 |
||||
| 60 Binder DK, Papadopoulos MC, Haggie PM, Verkman AS: In vivo measurement of brain extracellular space diffusion by cortical surface photobleaching. J Neurosci 2004;24:8049-8056. https://doi.org/10.1523/JNEUROSCI.2294-04.2004 |
||||
| 61 Haj-Yasein NN, Jensen V, Ostby I, Omholt SW, Voipio J, Kaila K, Petter Ottersen O, Hvalby O, Nagelhus EA: Aquaporin-4 regulates extracellular space volume dynamics during high-frequency synaptic stimulation:
A gene deletion study in mouse hippocampus. Glia 2012;60:867-874. https://doi.org/10.1002/glia.22319 |
||||
| 62 Mack AF, Wolburg H: A Novel Look at Astrocytes: Aquaporins, Ionic Homeostasis, and the Role of the Microenvironment for Regeneration in the CNS. Neuroscientist 2013;19:195-207. https://doi.org/10.1177/1073858412447981 |
||||
| 63 Xu Z, Xiao N, Chen Y, Huang H, Marshall C, Gao J, Xiao M: Deletion of aquaporin-4 in APP/PS1 mice exacerbates brain Aβ accumulation and memory deficits. Mol Neurodegener 2015;10:58. https://doi.org/10.1186/s13024-015-0056-1 |
||||
| 64 Skjolding AD, Rowland IJ, Søgaard LV, Praetorius J, Penkowa M, Juhler M: Hydrocephalus induces dynamic spatiotemporal regulation of aquaporin-4 expression in the rat brain. Cerebrospinal Fluid Res 2010;7:20. https://doi.org/10.1186/1743-8454-7-20 |
||||
| 65 Skjolding AD, Holst AV, Broholm H, Laursen H, Juhler M: Differences in distribution and regulation of astrocytic aquaporin‐4 in human and rat hydrocephalic brain. Neuropath Appl Neurobiol 2013;39:179-191. https://doi.org/10.1111/j.1365-2990.2012.01275.x |
||||
| 66 Ryskamp DA, Jo AO, Phuong TT, Verkman AS, Yarishkin O, MacAulay N, Križaj D: TRPV4 and AQP4 Channels Synergistically Regulate Cell Volume and Calcium Homeostasis in Retinal Müller Glia. J Neurosci 2015;35:13525-13537. https://doi.org/10.1523/JNEUROSCI.1987-15.2015 |
||||
| 67 Risher WC, Andrew RD, Kirov SA: Real-time passive volume responses of astrocytes to acute osmotic and ischemic stress in cortical slices and in vivo revealed by two-photon microscopy. Glia 2009;57:207-221. https://doi.org/10.1002/glia.20747 |
||||
| 68 Manley GT, Fujimura M, Ma T, Noshita N, Filiz F, Bollen AW, Chan P, Verkman AS: Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat Med 2000;6:159-163. https://doi.org/10.1038/72256 |
||||
| 69 Thrane AS, Rappold PM, Fujita T, Torres A, Bekar LK, Takano T, Peng W, Wang F, Thrane VR, Enger R, Haj-Yasein NN, Skare O, Holen T, Klungland A, Otterson OP, Nedergaard M, Nagelhus EA: Critical role of aquaporin-4 (AQP4) in astrocytic Ca2+ signaling events elicited by cerebral edema. Proc Natl Acad Sci U S A 2011;108:846-851. https://doi.org/10.1073/pnas.1015217108 |
||||
| 70 Tang G, Yang GY: Aquaporin-4: A Potential Therapeutic Target for Cerebral Edema. Int J Mol Sci 2016;17:1413. https://doi.org/10.3390/ijms17101413 |
||||
| 71 Papadopoulos MC, Manley GT, Krishna S, Verkman AS: Aquaporin-4 facilitates reabsorption of excess fluid in vasogenic brain edema. FASEB J 2004;18:1291-1293. https://doi.org/10.1096/fj.04-1723fje |
||||
| 72 Kitchen P, Day RE, Taylor LH, Salman MM, Bill RM, Conner MT, Conner AC: Identification and molecular mechanisms of the rapid tonicity-induced relocalization of the aquaporin 4 channel. J Biol Chem 2015;290:16873-16881. https://doi.org/10.1074/jbc.M115.646034 |
||||
| 73 Salman MM, Kitchen P, Woodroofe MN, Brown JE, Bill RM, Conner AC, Conner MT: Hypothermia increases aquaporin 4 (AQP4) plasma membrane abundance in human primary cortical astrocytes via a calcium/transient receptor potential vanilloid 4 (TRPV4)- and calmodulin-mediated mechanism. Eur J Neurosci 2017;46:2542-2547. https://doi.org/10.1111/ejn.13723 |
||||
| 74 Kitchen P, Salman MM, Halsey AM, Clarke-Bland C, MacDonald JA, Ishida H, Vogel HJ, Almutiri S, Logan A, Kreida S, Al-Jubair T, Winkel Missel J, Gourdon P, Törnroth-Horsefield S, Conner MT, Ahmed Z, Conner AC, Bill RM: Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020;181:784-799.e19. https://doi.org/10.1016/j.cell.2020.03.037 |
||||
| 75 Huber VJ, Tsujita M, Yamazaki M, Sakimura K, Nakada T: Identification of arylsulfonamides as Aquaporin 4 inhibitors. Bioorg Med Chem Lett 2007;17:1270-1273. https://doi.org/10.1016/j.bmcl.2006.12.010 |
||||
| 76 Huber VJ, Tsujita M, Kwee IL, Nakada T: Inhibition of aquaporin 4 by antiepileptic drugs. Bioorg Med Chem 2009;17:418-424. https://doi.org/10.1016/j.bmc.2007.12.038 |
||||
| 77 Huber VJ, Tsujita M, Nakada T: Identification of aquaporin 4 inhibitors using in vitro and in silico methods. Bioorg Med Chem 2009;17:411-417. https://doi.org/10.1016/j.bmc.2007.12.040 |
||||
| 78 Igarashi H, Huber VJ, Tsujita M, Nakada T: Pretreatment with a novel aquaporin 4 inhibitor, TGN-020, significantly reduces ischemic cerebral edema. Neurol Sci 2011;32:113-116. https://doi.org/10.1007/s10072-010-0431-1 |
||||
| 79 Li J, Jia Z, Xu W, Guo W, Zhang M, Bi J, Cao Y, Fan Z, Li G: TGN-020 alleviates edema and inhibits astrocyte activation and glial scar formation after spinal cord compression injury in rats. Life Sci 2019;222:148-157. https://doi.org/10.1016/j.lfs.2019.03.007 |
||||
| 80 Stokum JA, Kwon MS, Woo SK, Tsymbalyuk O, Vennekens R, Gerzanich V, Simard JM: SUR1-TRPM4 and AQP4 form a heteromultimeric complex that amplifies ion/water osmotic coupling and drives astrocyte swelling. Glia 2018;66:108-125. https://doi.org/10.1002/glia.23231 |
||||
| 81 Illarionova NB, Gunnarson E, Li Y, Brismar H, Bondar A, Zelenin S, Aperia A: Functional and molecular interactions between aquaporins and Na,K-ATPase. Neurosci 2010;168:915-925. https://doi.org/10.1016/j.neuroscience.2009.11.062 |
||||
| 82 Mutai H, Heller S: Vertebrate and invertebrate TRPV-like mechanoreceptors. Cell Calcium 2003;33:471-478. https://doi.org/10.1016/S0143-4160(03)00062-9 |
||||
| 83 Liedtke W: Transient receptor potential vanilloid channels functioning in transduction of osmotic stimuli.
J Endocrinol 2006;191:515-523. https://doi.org/10.1677/joe.1.07000 |
||||
| 84 Becker D, Blase C, Bereiter-Hahn J, Jendrach M: TRPV4 exhibits a functional role in cell-volume regulation.
J Cell Sci 2005;118:2435-2440. https://doi.org/10.1242/jcs.02372 |
||||
| 85 Liedtke W, Friedman JM: Abnormal osmotic regulation in trpv4-/- mice. Proc Natl Acad Sci U S A 2003;100:13698-13703. https://doi.org/10.1073/pnas.1735416100 |
||||
| 86 Benfenati V, Amiry-Moghaddam M, Caprini M, Mylonakou MN, Rapisarda C, Petter Ottersen O, Ferroni S: Expression and functional characterization of transient receptor potential vanilloid-related channel 4 (TRPV4) in rat cortical astrocytes. Neuroscience 2007;148:876-892. https://doi.org/10.1016/j.neuroscience.2007.06.039 |
||||
| 87 Becker D, Bereiter-Hahn J, Jendrach M: Functional interaction of the cation channel transient receptor potential vanilloid 4 (TRPV4) and actin in volume regulation. Eur J Cell Biol 2009;88:141-152. https://doi.org/10.1016/j.ejcb.2008.10.002 |
||||
| 88 Vincent F, Duncton MA: TRPV4 agonists and antagonists. Curr Top Med Chem 2011;11:2216-2226. https://doi.org/10.2174/156802611796904861 |
||||
| 89 Wei ZL, Nguyen MT, O'Mahony DJ, Acevedo A, Zipfel S, Zhang Q, Liu L, Dourado M, Chi C, Yip V, DeFalco J, Gustafson A, Emerling DE, Kelly MG, Kincaid J, Vincent F, Duncton MA: Identification of orally-bioavailable antagonists of the TRPV4 ion-channel. Bioorg Med Chem Lett 2015;25:4011-4015. https://doi.org/10.1016/j.bmcl.2015.06.098 |
||||
| 90 Goyal N, Skrdla P, Schroyer R, Kumar S, Fernando D, Oughton A, Norton N, Sprecher DL, Cheriyan J: Clinical Pharmacokinetics, Safety, and Tolerability of a Novel, First-in-Class TRPV4 Ion Channel Inhibitor, GSK2798745, in Healthy and Heart Failure Subjects. Am J Cardiovasc Drugs 2019;19:335-342. https://doi.org/10.1007/s40256-018-00320-6 |
||||
| 91 Tanaka K, Matsumoto S, Yamada T, Yamasaki R, Suzuki M, Kido MA, Kira JI: Reduced Post-ischemic Brain Injury in Transient Receptor Potential Vanilloid 4 Knockout Mice. Front Neurosci 2020;14:453. https://doi.org/10.3389/fnins.2020.00453 |
||||
| 92 Shen J, Tu L, Chen T, Tan T, Wang Y, Wang S: TRPV4 channels modulate Ca2+-induced Ca2+ release in mouse neurons and trigger endoplasmic reticulum stress after intracerebral hemorrhage. Brain Res Bull 2019;146:143-152. https://doi.org/10.1016/j.brainresbull.2018.11.024 |
||||
| 93 Jie P, Lu Z, Hong Z, Li L, Zhou L, Li Y, Zhou R, Zhou Y, Du Y, Chen L, Chen L: Activation of Transient Receptor Potential Vanilloid 4 is Involved in Neuronal Injury in Middle Cerebral Artery Occlusion in Mice. Mol Neurobiol 2016;53:8-17. https://doi.org/10.1007/s12035-014-8992-2 |
||||
| 94 Ryskamp DA, Jo AO, Frye AM, Vazquez-Chona F, MacAulay N, Thoreson WB, Križaj D: Swelling and eicosanoid metabolites differentially gate TRPV4 channels in retinal neurons and glia. J Neurosci 2014;34:15689-15700. https://doi.org/10.1523/JNEUROSCI.2540-14.2014 |
||||
| 95 Asao Y, Tobori S, Kakae M, Nagayasu K, Shibasaki K, Shirakawa H, Kaneko S: Transient receptor potential vanilloid 4 agonist GSK1016790A improves neurological outcomes after intracerebral hemorrhage in mice. Bioche Biophys Res Commun 2020;529:590-595. https://doi.org/10.1016/j.bbrc.2020.06.103 |
||||
| 96 Chen CK, Hsu PY, Wand TM, Miao ZF, Lin RT, Juo SHH: TRPV4 activation contributes functional recovery from ischemic stroke via angiogenesis and neurogenesis. Mol Neurobiol 2018;55:4127-4135. https://doi.org/10.1007/s12035-017-0625-0 |
||||
| 97 Liu M, Liu X, Wang L, Wang Y, Dong F, Wu J, Qu X, Liu Y, Liu Z, Fan H, Yao R: TRPV4 Inhibition Improved Myelination and Reduced Glia Reactivity and Inflammation in a Cuprizone-Induced Mouse Model of Demyelination. Front Cell Neurosci 2018;12:392. https://doi.org/10.3389/fncel.2018.00392 |
||||
| 98 Hochstetler AE, Smith HM, Preston DC, Reed MM, Territo PR, Shim JW, Fulkerson D, Blazer-Yost BL: TRPV4 antagonists ameliorate ventriculomegaly in a rat model of hydrocephalus. JCI Insight 2020;5:e137646. https://doi.org/10.1172/jci.insight.137646 |
||||
| 99 Chen M, Simard JM: Cell swelling and a nonselective cation channel regulated by internal Ca2+ and ATP in native reactive astrocytes from adult rat brain. J Neurosci 2001;21:6512-6521. https://doi.org/10.1523/JNEUROSCI.21-17-06512.2001 |
||||
| 100 Chen M, Dong Y, Simard JM: Functional coupling between sulfonylurea receptor type 1 and a nonselective cation channel in reactive astrocytes from adult rat brain. J Neurosci 2003;23:8568-8577. https://doi.org/10.1523/JNEUROSCI.23-24-08568.2003 |
||||
| 101 Simard JM, Chen M, Tarasov KV, Bhatta S, Ivanova S, Melnitchenko L, Tsymbalyuk N, West GA, Gerzanich V: Newly expressed SUR1-regulated NC(Ca-ATP) channel mediates cerebral edema after ischemic stroke. Nat Med 2006;12:433-440. https://doi.org/10.1038/nm1390 |
||||
| 102 Mehta RI, Tosun C, Ivanova S, Tsymbalyuk N, Famakin BM, Kwon MS, Castellani RJ, Gerzanich V, Simard JM: Sur1-Trpm4 Cation Channel Expression in Human Cerebral Infarcts. J Neuropathol Exp Neurol 2015;74:835-849. https://doi.org/10.1097/NEN.0000000000000223 |
||||
| 103 Sheth KN, Simard JM, Elm J, Kronenberg G, Kunte H, Kimberly WT: Human data supporting glyburide in ischemic stroke. Acta Neurochir Suppl 2016;121:13-18. https://doi.org/10.1007/978-3-319-18497-5_3 |
||||
| 104 Delalande C, Awale M, Rubin M, Probst D, Ozhathil LC, Abriel H, Reymond JL: Optimizing TRPM4 inhibitors in the MHFP6 chemical space. Eur J Med Chem 2019;166:167-177. https://doi.org/10.1016/j.ejmech.2019.01.048 |
||||
| 105 Chen B, Gao Y, Wei S, Low SW, Ng G, Yu D, Tu TM, Soong TW, Nilius B, Liao P: TRPM4-specific blocking antibody attenuates reperfusion injury in a rat model of stroke. Pflugers Arch 2019;471:1455-1466. https://doi.org/10.1007/s00424-019-02326-8 |
||||
| 106 Walch E, Murphy TR, Cuvelier N, Aldoghmi M, Morozova C, Donohue J, Young G, Samant A, Garcia S, Alvarez C, Bilas A, Davila D, Binder DK, Fiacco TA: Astrocyte-Selective Volume Increase in Elevated Extracellular Potassium Conditions Is Mediated by the Na+/K+ ATPase and Occurs Independently of Aquaporin 4. ASN Neuro 2020;12:1759091420967152. https://doi.org/10.1177/1759091420967152 |
||||
| 107 Murakami S, Kurachi S: Mechanisms of astrocytic K+ clearance and swelling under extracellular K+ concentrations. J Physiol Sci 2016;66:127-142. https://doi.org/10.1007/s12576-015-0404-5 |
||||
| 108 Chen H, Sun D: The role of Na-K-Cl co-transporter in cerebral ischemia. Neurol Res 2005;27:280-286. https://doi.org/10.1179/016164105X25243 |
||||
| 109 Staub F, Stoffel M, Berger S, Eriskat J, Baethmann A: Treatment of vasogenic brain edema with the novel Clᴄtransport inhibitor torasemide. J Neurotrauma 1994;11:679-690. https://doi.org/10.1089/neu.1994.11.679 |
||||
| 110 MacVicar B, Hochman D: Imaging of synaptically evoked intrinsic optical signals in hippocampal slices.
J Neurosci 1991;11:1458-1469. https://doi.org/10.1523/JNEUROSCI.11-05-01458.1991 |
||||
| 111 MacVicar BA, Feighan D, Brown A, Ransom B: Intrinsic optical signals in the rat optic nerve: role for K(+) uptake via NKCC1 and swelling of astrocytes. Glia 2002;37:114-123. https://doi.org/10.1002/glia.10023 |
||||
| 112 Su G, Kintner DB, Flagella M, Shull GE, Sun D: Astrocytes from Na(+)-K(+)-Cl(−) cotransporter-null mice exhibit absence of swelling and decrease in EAA release. Am J Physiol Cell Physiol 2002;282:C1147-C1160. https://doi.org/10.1152/ajpcell.00538.2001 |
||||
| 113 Jayakumar AR, Liu M, Moriyama M, Ramakrishnan R, Forbush B 3rd, Reddy PV, Norenberg MD: Na-K-Cl Cotransporter-1 in the mechanism of ammonia-induced astrocyte swelling. J Biol Chem 2008;283:33874-33882. https://doi.org/10.1074/jbc.M804016200 |
||||
| 114 Zeuthen T, MacAulay N: Cotransport of water by Na+-K+-2Cl- cotransporters expressed in Xenopus oocytes: NKCC1 versus NKCC2. J Physiol 2012;590:1139-1154. https://doi.org/10.1113/jphysiol.2011.226316 |
||||
| 115 Zhang M, Cui Z, Cui H, Cao Y, Zhong C, Wang Y: Astaxanthin alleviates cerebral edema by modulating NKCC1 and AQP4 expression after traumatic brain injury in mice. BMC Neurosci 2016;17:60. https://doi.org/10.1186/s12868-016-0295-2 |
||||
| 116 Parkerson KA, Sontheimer H: Contribution of chloride channels to volume regulation of cortical astrocytes. Am J Physiol Cell Physiol 2003;284:C1460-C1467. https://doi.org/10.1152/ajpcell.00603.2002 |
||||
| 117 Parkerson KA, Sontheimer H: Biophysical and pharmacological characterization of hypotonically activated chloride currents in cortical astrocytes. Glia 2004;46:419-436. https://doi.org/10.1002/glia.10361 |
||||
| 118 Hyzinski-Garcia MC, Rudkouskaya A, Mongin AA: LRRC8A protein is indispensable for swelling-activated and ATP-induced release of excitatory amino acids in rat astrocytes. J Physiol 2014;592:4855-4862. https://doi.org/10.1113/jphysiol.2014.278887 |
||||
| 119 Formaggio F, Saracino E, Mola MG, Rao SB, Amiry-Moghaddam M, Muccini M, Zamboni R, Nicchia GP, Caprini M, Benfenati V: LRRC8A is essential for swelling-activated chloride current and for regulatory volume decrease in astrocytes. FASEB J 2019;33:101-113. https://doi.org/10.1096/fj.201701397RR |
||||
| 120 Murphy TR, Binder DK, Fiacco TA: Turning down the volume: Astrocyte volume change in the generation and termination of epileptic seizures. Neurobiol Dis 2017;104:24-32. https://doi.org/10.1016/j.nbd.2017.04.016 |
||||
| 121 Qiu Z, Dubin AE, Mathur J, Tu B, Reddy K, Miraglia LJ, Reinhardt J, Orth AP, Patapoutian A: SWELL1, a plasma membrane protein, is an essential component of volume-regulated anion channel. Cell 2014;157:447-458. https://doi.org/10.1016/j.cell.2014.03.024 |
||||
| 122 Voss FK, Ullrich F, Munch J, Lazarow K, Lutter D, Mah N, Andrade-Navarro MA, von Kries JP, Stauber T, Jentsch TJ: Identification of LRRC8 heteromers as an essential component of the volume-regulated anion channel VRAC. Science 2014;344:634-638. https://doi.org/10.1126/science.1252826 |
||||
| 123 Kimelberg HK, Rutledge E, Goderie S, Charniga C: Astrocytic swelling due to hypotonic or high K+ medium causes inhibition of glutamate and aspartate uptake and increases their release. J Cereb Blood Flow Metab 1995;15:409-416. https://doi.org/10.1038/jcbfm.1995.51 |
||||
| 124 Vitarella D, DiRisio DJ, Kimelberg HK, and Aschner M: Potassium and taurine release are highly correlated with regulatory volume decrease in neonatal primary rat astrocyte cultures. J Neurochem 1994;63:1143-1149. https://doi.org/10.1046/j.1471-4159.1994.63031143.x |
||||
| 125 Olson JE, Li GZ: Increased potassium, chloride, and taurine conductances in astrocytes during hypoosmotic swelling. Glia 1997;20:254-261. https://doi.org/10.1002/(SICI)1098-1136(199707)20:3<254::AID-GLIA9>3.0.CO;2-7 |
||||
| 126 Benfenati V, Nicchia GP, Svelto M, Rapisarda C, Frigeri A, Ferroni S: Functional down-regulation of volume-regulated anion channels in AQP4 knockdown cultured rat cortical astrocytes. J Neurochem 2007;100:87-104. https://doi.org/10.1111/j.1471-4159.2006.04164.x |
||||
| 127 Orkand R, Nicholls J, Kuffler S: Effect of nerve impulses on the membrane potential of glial cells in the central nervous system of amphibia. J Neurophysiol 1966;29:788-806. https://doi.org/10.1152/jn.1966.29.4.788 |
||||
| 128 Howe MW, Feig SL, Osting SM, Haberly LB: Cellular and subcellular localization of Kir2.1 subunits in neurons and glia in piriform cortex with implications for K+ spatial buffering. J Comp Neurol 2008;506:877-893. https://doi.org/10.1002/cne.21534 |
||||
| 129 Kang SJ, Cho SH, Park K, Yi J, Yoo SJ, Shin KS: Expression of Kir2.1 channels in astrocytes under pathophysiological conditions. Mol Cells 2008;25:124-130. | ||||
| 130 Nagelhus EA, Mathiisen TM, Petter Ottersen O: Aquaporin-4 in the central nervous system: cellular and subcellular distribution and coexpression with KIR4.1. Neurosci 2004;129:905-913. https://doi.org/10.1016/j.neuroscience.2004.08.053 |
||||
| 131 Francis C, Natarajan S, Lee MT, Khaladkar M, Buckley PT, Sul JY, Eberwine J, Kim J: Divergence of RNA localization between rat and mouse neurons reveals the potential for rapid brain evolution. BMC Genomics 2014;15:883. https://doi.org/10.1186/1471-2164-15-883 |
||||
| 132 Xu X, Stoyanova EI, Lemiesz AE, Xing J, Mash DC, Heintz N: Species and cell-type properties of classically defined human and rodent neurons and glia. eLife 2018;7:e37551. https://doi.org/10.7554/eLife.37551 |
||||
| 133 Xu H, Fu Y, Tian W, Cohen DM: Glycosylation of the osmoresponsive transient receptor potential channel TRPV4 on Asn-651 influences membrane trafficking. Am J Physiol Renal Physiol 2006;290:F1103-F1109. https://doi.org/10.1152/ajprenal.00245.2005 |
||||
| 134 Semple BD, Blomgren K, Gimlin K, Ferriero DM, Noble-Haeusslein LJ: Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog Neurobiol 2013;106-107:1-16. https://doi.org/10.1016/j.pneurobio.2013.04.001 |
||||
| 135 Xu H, Fame RM, Sadegh C, Sutin J, Naranjo C, Syau D, Cui J, Shipley FB, Vernon A, Gao F, Zhang Y, Holtzman MJ, Heiman M, Warf BC, Lin P, Lehtinen MK: Choroid plexus NKCC1 mediates cerebrospinal fluid clearance during mouse early postnatal development. Nat Commun 2021;12:447. https://doi.org/10.1038/s41467-020-20666-3 |
||||