×
![]()
Corresponding Author: Dr. Razieh Mohammad Jafari and Prof. Ahmad Reza Dehpour
Experimental Medicine Research Center, Tehran University of Medical Sciences, Tehran 13145-784 (Iran)
Tel. +98 21 88973652, Fax +98 21 66402569, E-Mail rmjafari@sina.tums.ac.ir; dehpoura@sina.tums.ac.ir; dehpour@yahoo.com
Sodium Selenite Modulates IDO1/Kynurenine, TLR4, NF-κB and Bcl2/Bax Pathway and Mitigates Acetic Acid-Induced Colitis in Rat
Moein Alaa,b Razieh Mohammad Jafaria Hossein Nematiana,b Amir Shadboorestanc Ahmad Reza Dehpoura,b
aExperimental Medicine Research Center, Tehran University of Medical Sciences, Tehran, Iran, bDepartment of Pharmacology, School of Medicine, Tehran University of Medical Sciences, Tehran, Iran, cDepartment of Toxicology, Faculty of Medical Sciences, Tarbiat Modares University, Tehran, Iran,
Introduction
Colitis is defined as inflammation of the colon and is a major component of IBD. It varies in depth (transmural or non-transmural) and length (pan colitis or segmental), so the treatment will vary, as well. A sick colon imposes many limitations in eating habits, affects bowel movement, produces abdominal discomfort, and disturbs the normal living and function [1, 2]. Contrary to consuming different immunosuppressive drugs such as corticosteroids, anti-TNF agents, 5-aminosalicylic acid derivatives and cytotoxic drugs, a large number of colitis cases particularly patients with IBD remain untreated [3, 4]. Except the failure rate, these drugs bring severe adverse effects, which is not always tolerated and some of them impose high economic burden.
Selenium is a micronutrient with antioxidant function. Selenium modulates inflammation by altering the expression of pro- and anti-inflammatory mediators [5, 6]. It is relatively safe but consuming very high doses of selenium leads to selenium toxicity that presents with diarrhea, nausea, fatigue, hair loss, arthralgia, and nail discoloration [7]. It was reported that serum selenium concentration of 400–30,000 μg/L and 500–1400 μg/L were associated with acute and chronic selenium toxicity, respectively [8]. These concentrations of selenium are much higher than normal serum concentrations of selenium (1.14 μmol/L or 90 μg/L) [9]. Selenium deficiency was associated with decreased selenoproteins expression, increased oxidative stress and inflammatory cytokines production in the animal models and selenium supplementation attenuated these inflammatory changes [10, 11]. Studies revealed that selenium and selenoproteins alter the expression of TLR4, NF-κB, bcl2 and bax to suppress inflammation and enhance cellular longevity [12-14].
Several clinical studies have reported that selenium deficiency is observed in a notable percentage of patients with IBD and may be a contributory factor for colitis [15-17]. Consistently, selenium deficiency expanded the extent of tissue destruction in animal models of colitis [18]. Similarly, it was found that selenium alone or in combination with other substances such as vitamin E mitigates the intensity of damage due to its anti-oxidant properties [19-21]. Furthermore, selenium supports gut commensal microbiota and enhances their protective effects on intestinal lining [21].
The aim of this study was to evaluate the effect of sodium selenite on mild, moderate and severe colitis at the gross and microscopic level. In addition, we investigated the effect of sodium selenite on the expression of inflammatory mediators and apoptosis regulators.
Materials and Methods
Animals
This experiment was performed with seventy-two male Wistar rats, aged 10 to 12 weeks and weighing 200 to 250 g. Animals were obtained from the Animal house of Faculty of medicine, Tehran University of Medical Sciences, Tehran, Iran. All animals were housed in a controlled ambient temperature (22±2 °C) on a 12:12h light/dark cycle. Rats were fed a standard diet and had free access to water. After acclimatization to their environment, the study began. Anesthesia was induced by ketamine (87 mg/kg) and xylazine (13 mg/kg) [22] and rats were followed until 1 hour after awaking. All efforts were spared to reduce animal suffering and the number of animals used. All animal experiments were approved by the Institutional Animal Care and Use Committee of Tehran University of Medical Sciences (IR.TUMS.VCR.REC.1397.833). The study was conducted in accordance with the Guidelines for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH publication, 8th Ed.).
Acute Colitis Model
Experimental rats fasted for at least 24 h before the induction of colitis with free access to water. Colonic inflammation was induced under light anesthesia by administering 2 ml of acetic acid in three increasing concentrations (4% - 6% - 8% v/v) dissolved in 0.9% physiologic saline. A polypropylene tube with an outer diameter of 3 mm was used for intra-rectal acid instillation. The tube was inserted intrarectally into the colon to a distance of 8 cm from the anus. Rats were hold vertically (with their head downward) for 2 minutes to prevent rapid outflow of acetic acid.
Treatment Schedule
After induction of acute colitis by acetic acid, treatment was performed with two consecutive gavages of 0.5 mg/kg sodium selenite on days 0 and 1. Control group received an equal amount of normal saline. Two days after the induction of colitis, the animals underwent colectomy surgery, extracting their terminal colon. This part was used for microscopic, macroscopic and molecular assessments (Fig. 1).

Animals Grouping
Experimental rats were randomly divided into 6 groups (n=6) as shown in Table 1.
Because of the high mortality rate, the sample size in groups 5 and 6 did not reach to the aim of this study. Therefore, the investigation was ended in these groups because of the ethics. Specimens from group 3 and 4 were used for molecular assessments.
Macroscopic and Microscopic Measurement
The terminal part of colon was opened longitudinally to expose the internal luminal surface. After washing mucosal surface and removing the feces, the photos were taken from the gross view. Then samples were collected for molecular and microscopic measurements.
For histopathological examination, specimens were fixed in 4% buffered paraformaldehyde solution (PFA 4%). Thereafter, sections with 5-μm thickness were cut and stained with hematoxylin and eosin (H&E). An experienced pathologist blinded to the identity of specimens, performed all histopathological processing and interpreted the results.
Measurement of Pro-Inflammatory Cytokines by ELISA
For molecular assessments specimens were snap frozen in -80 °C and kept in -20 °C before processing. The colon tissue samples were sonicated (10 w, 2×5s) in a mixture containing protease inhibitors and then centrifuged at 50,000×g for 20 min at 4 °C. TNF-α (DuoSet®, DY510-05), INF-γ (DuoSet®, Rat IFN-γ DY585) and Myeloperoxidase (MPO Colorimetric Activity Assay Kit MAK068), MDA (Teb Pazhouhan Razi (TRP), Tehran, Iran), IL22 (ERA27RB), IL17 (DY4437) and kynurenine (Rat Kynurenine (KYNU) Elisa kit MBS745507) were measured in the supernatant by ELISA as explained by the manufacturer.
Western Blot
Colon tissue was sonicated and homogenized in lysis buffer (included: Tris HCl, SDS, DTT, Glycerol, NP40). The homogenate was centrifuged at 10000 g for 10 min in 4 °C. The supernatants were extracted for immunoblotting protein expression. After the samples were boiled at 100 °C for 5 minutes, loading samples on sodium dodecyl sulfate-polyacrylamide gels (SDS-PAGE) and using running buffer, transferred to Polyvinylidene difluoride (PVDF) membranes. The nonspecific binding sites on membranes were blocked 90 min with 5% non-fat skim milk at room temperature and incubated with the following primary antibodies overnight: IDO1 (Santa Cruz Biotechnology, sc-137012), p-NF-κB p65 (Santa Cruz Biotechnology, sc-136548), Anti-NF-κB p65 antibody (ab16502, abcam), Bax (Santa Cruz Biotechnology, sc-7480), TLR4 (Santa Cruz Biotechnology, sc-293072), Bcl2 (Santa Cruz Biotechnology, sc-492), and β-actin (Santa Cruz Biotechnology, sc-47778). Membranes were then washed with TBST (TBS+ tween 80) and incubated with secondary antibodies for 60 min at room temperature. BM chemiluminescence western blot kit used for detection immunostained protein bands by gel doc. An open source image processing program, Image J, was applied to quantify the optical density of each band. β-actin was used as an internal control. The relative activity of all proteins was calculated by Prism 7 (GraphPad Inc.) and compared with the control group [23].
Statistical Analysis
Data are presented as Mean ± SEM and analyzed using GraphPad Prism7 software (version 6.07). Tests of homogeneity of variance were used to ensure normal distribution of data. One-way analysis of variance (ANOVA) and T test were used to analyze the data. Probability (p) value less than 0.05 was considered significant.
Results
Macroscopic Finding
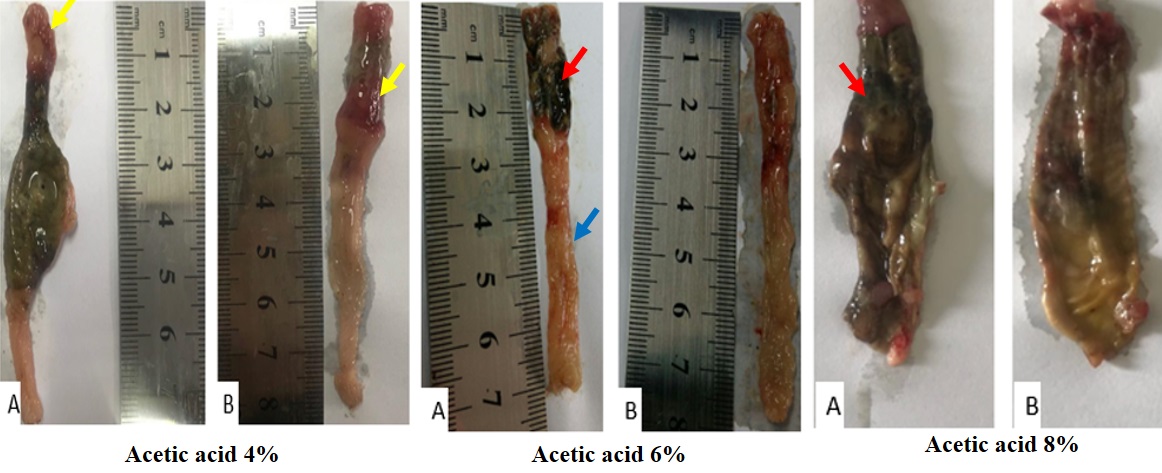
Acetic acid enema led to colitis in all rats. Gross colonic injuries characterized by diffuse mucosal edema and granularity (blue arrow), erythema (yellow arrow), ulceration and even necrosis (red arrow) were observed in saline-treated group. The intensity and extent of destruction increased proportional to the increase of acid concentration. Oral administration of sodium selenite after induction of colitis alleviated the macroscopic signs of destruction. Treatment with sodium selenite reduced the extent of the colonic injury and mitigated the degree of edema, granulation and erythema. In addition, differences were more perceptible in lower concentrations of acetic acid (Fig. 2).
Histopathological Findings
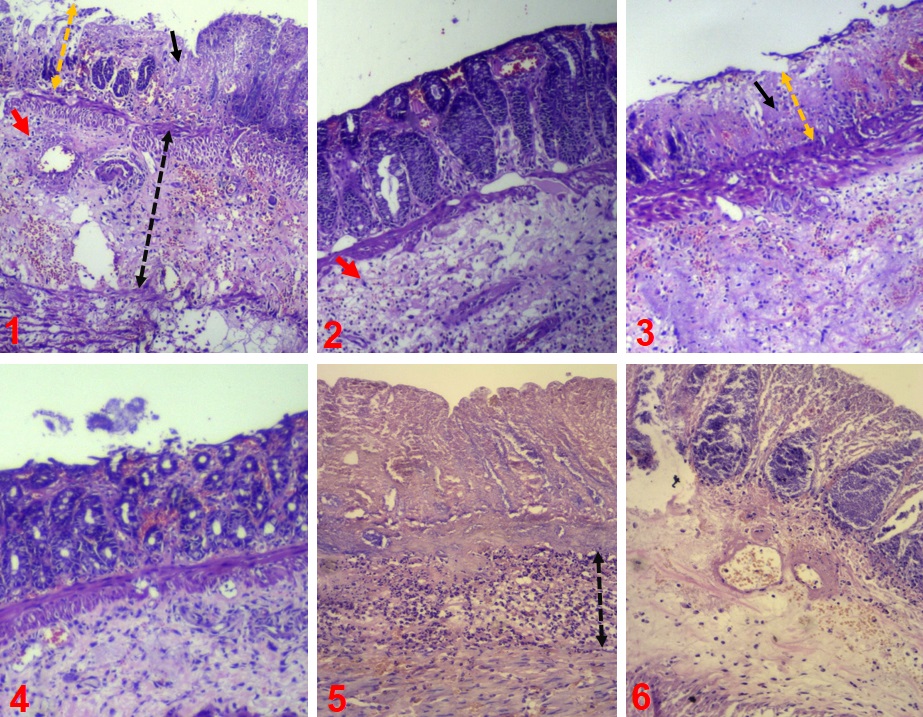
In saline-treated groups, a significant infiltration of inflammatory cells was observed. Inflammation of crypts (cryptitis), collection of neutrophils within crypts lumen (crypt abscess) and mucosal ulcers were prominent in the saline-treated groups compared to sodium selenite-treated groups. Histopathological analysis of the colon revealed that acetic acid-induced changes of colon tissue such as loss of tissue architecture, cellular infiltration into the mucosa and submucosa, abscess formation, hemorrhage in the crypts were alleviated in animals treated with sodium selenite (Fig. 3 & Fig. 4).
Sodium Selenite Exhibits Anti-Oxidant Effects in the Treatment of Acetic Acid-Induced Colitis
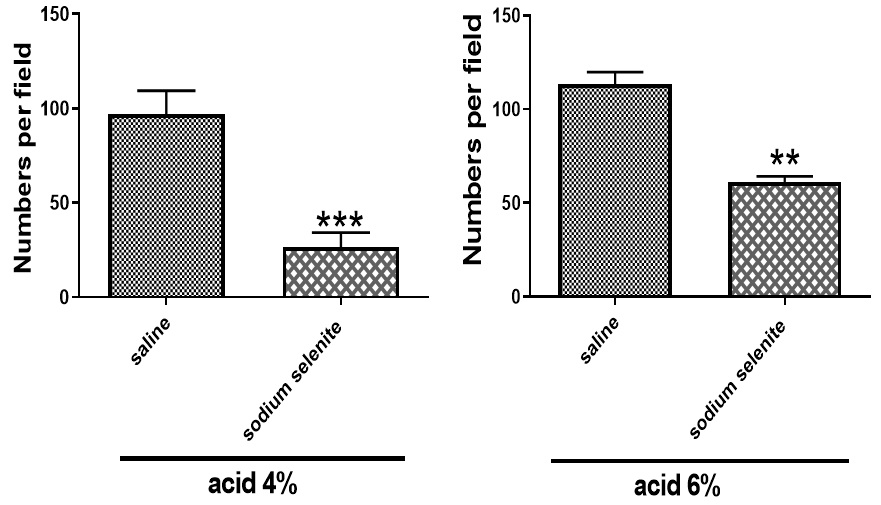
MDA assay kit (Teb Pazhouhan Razi, Iran, Code No. TPR-MDA-96T) was used for measurement of MDA as a marker of oxidative stress. As shown in Fig. 4, MDA levels were markedly decreased in acetic acid 4% (**p<0.01) and acetic acid 6% (*p<0.05), compared with saline-treated groups. MPO activity was significantly (*p<0.05) lower in sodium selenite-treated group (Fig. 5).
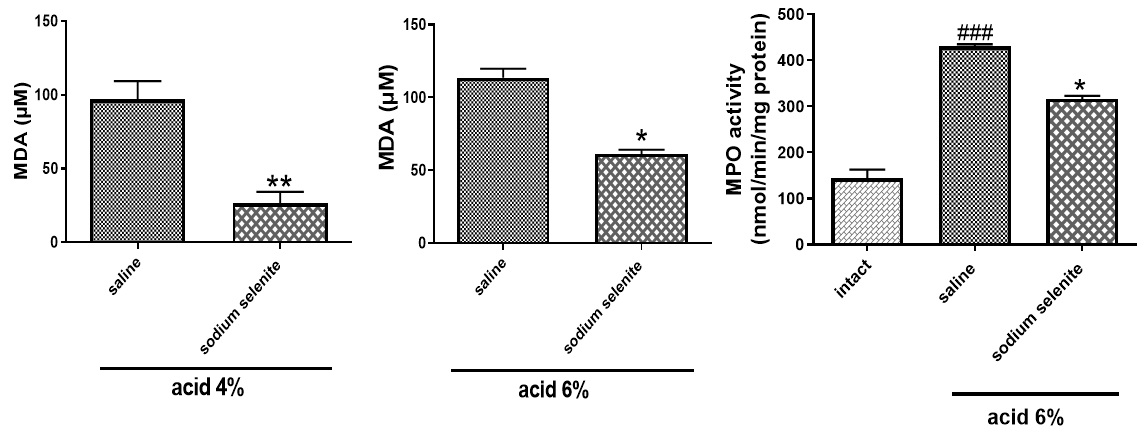
Sodium Selenite Exhibits Anti-inflammatory Effects in the Treatment of Acetic Acid-Induced Colitis
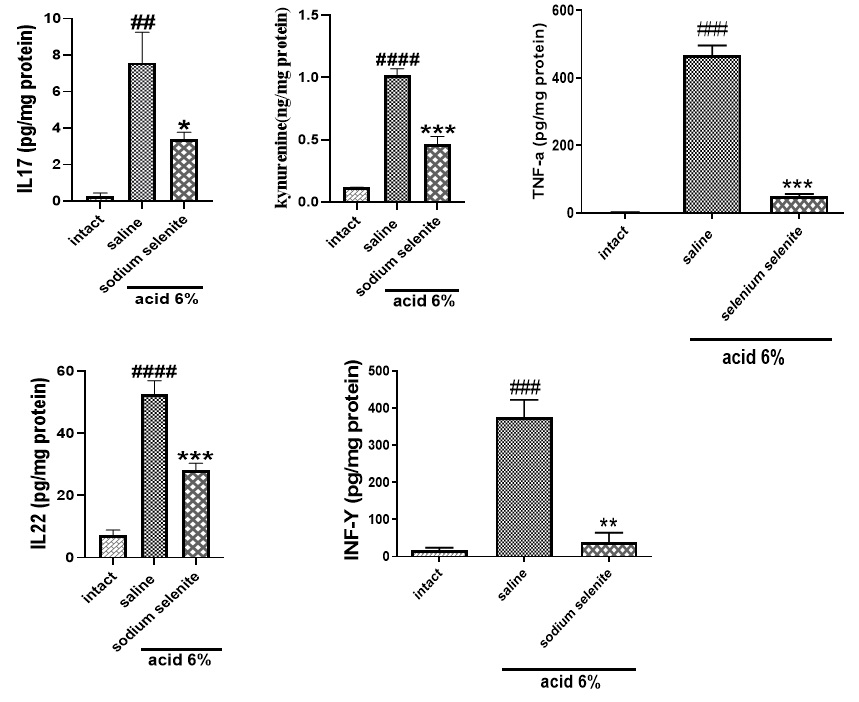
As shown in Fig. 5, treatment with sodium selenite significantly decreased TNF-α (***p<0.001) and INF-γ (**p<0.01) levels. In addition, sodium selenite could significantly decrease the increased levels of IL22 (***p<0.001), IL17 (*p<0.05) and kynurenine (***p<0.001) (Fig. 6).
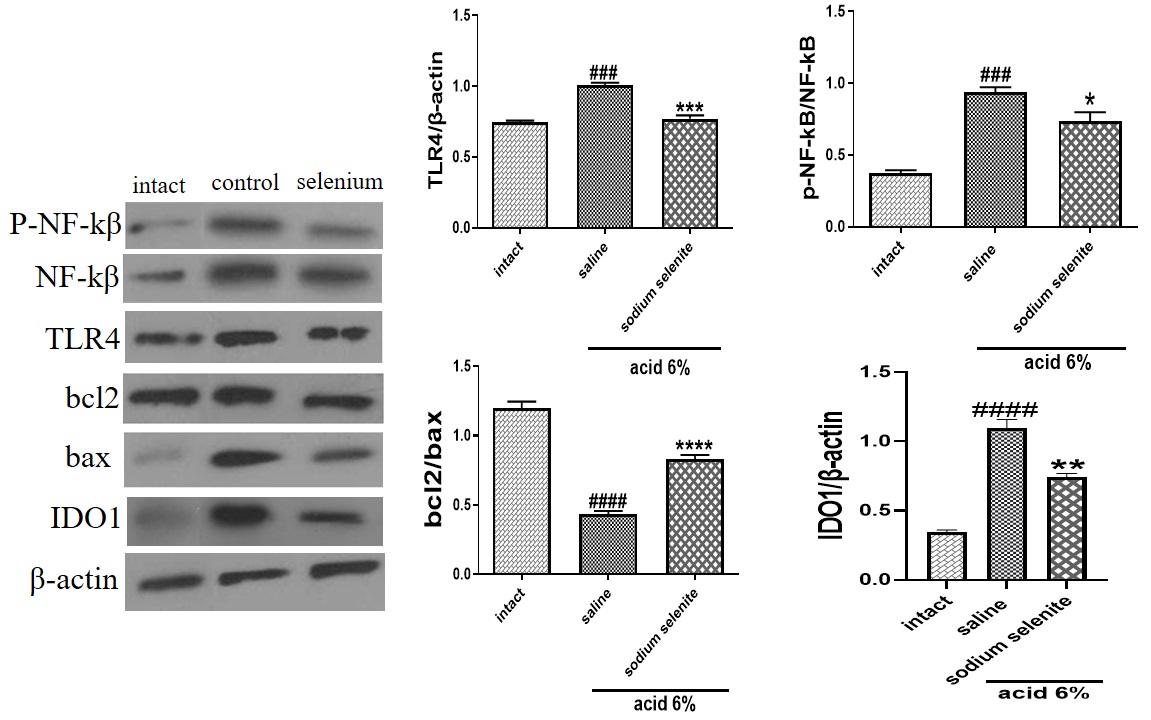
Sodium Selenite Modulate TLR4, NF-κB, IDO1 and Apoptosis Signaling Pathways to Protect against Acetic Acid-Induced Colitis in Rats
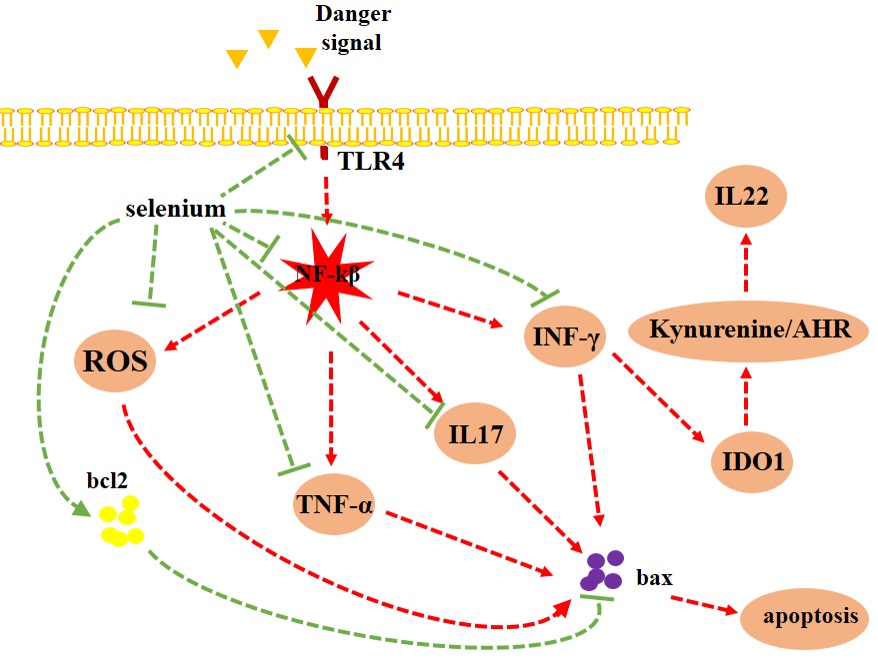
Western blotting was used to measure major inflammatory signaling pathways and apoptosis markers after induction and treatment of colitis. Acid enema caused a significant increase in TLR4/β-actin (###p<.0.001), p-NF-κB/NF-κB (###p<0.001) and indoleamine 2,3-dioxygenase IDO1/β-actin (####p<0.0001). In addition, acid enema significantly decreased bcl2/Bax (####p<0.0001). Treatment with sodium selenite 0.5 mg/kg meaningfully decreased TLR4 (***p<0.001), p-NF-κB/NF-κB (*p<0.05) and IDO1/β-actin (**p<0.01) and increased bcl2//Bax ratio (****p<0.0001) (Fig. 7).
Author Contributions
In this study every member respectively performed these responsibilities:
Moein Ala (surgery and following animals, writing the article), Razieh Mohammad Jafari (Western blot performance and editing assay, co-corresponding), Hossein Nematian (surgery), Amir Shadboorestan (Western blot performance), Ahmad Reza Dehpour (conceived and designed research, principal investigator).
Funding
This work was supported by a grant from Experimental Medicine Research Center, Tehran University of Medical Sciences, Tehran, Iran (Grant No. 97-03-158-40561) and Iran National Science Foundation (INSF) (Grant No. 96002757).
The authors declare that no conflicts of interest exist.
| 1 Heikenen JB, Werlin SL, Brown CW, Balint JP: Presenting symptoms and diagnostic lag in children with inflammatory bowel disease. Inflamm Bowel Dis 1999;5:158-160. https://doi.org/10.1097/00054725-199908000-00002 |
||||
| 2 Spray C, Debelle G, Murphy M: Current diagnosis, management and morbidity in paediatric inflammatory bowel disease. Acta Paediatr 2001;90:400-405. https://doi.org/10.1080/08035250118892 |
||||
| 3 Yanai H, Hanauer SB: Assessing response and loss of response to biological therapies in IBD. Am J Gastroenterol 2011;106:685. https://doi.org/10.1038/ajg.2011.103 |
||||
| 4 Kim J, Ham N, Oh E, Song E, Youn E, Kwon E, Cho Y, Hwang S, Park S, Yang D: P277 Real life effectiveness and safety of vedolizumab induction and maintenance therapy for Korean IBD patients in whom anti-TNF treatment failed: a prospective cohort study. J Crohns Colitis 2019;13:S237-S237. https://doi.org/10.1093/ecco-jcc/jjy222.401 |
||||
| 5 Hughes D, Kunická T, Schomburg L, Liška V, Swan N, Souček P: Expression of Selenoprotein Genes and Association with Selenium Status in Colorectal Adenoma and Colorectal Cancer. Nutrients 2018;10:1812. https://doi.org/10.3390/nu10111812 |
||||
| 6 Zhang JL, Xu B, Huang XD, Gao YH, Chen Y, Shan AS: Selenium deficiency affects the mRNA expression of inflammatory factors and selenoprotein genes in the kidneys of broiler chicks. Biol Trace Elem Res 2016;171:201-207. https://doi.org/10.1007/s12011-015-0512-3 |
||||
| 7 MacFarquhar JK, Broussard DL, Melstrom P, Hutchinson R, Wolkin A, Martin C, Burk RF, Dunn JR, Green AL, Hammond R: Acute selenium toxicity associated with a dietary supplement. Arch Intern Med 2010;170:256-261. https://doi.org/10.1001/archinternmed.2009.495 |
||||
| 8 Nuttall KL: Evaluating selenium poisoning. Ann Clin Lab Sci 2006;36:409-420. | ||||
| 9 Yang KC, Lee LT, Lee YS, Huang HY, Chen CY, Huang KC: Serum selenium concentration is associated with metabolic factors in the elderly: a cross-sectional study. Nutr Metab (Lond) 2010;7:1-7. https://doi.org/10.1186/1743-7075-7-38 |
||||
| 10 Liu S, Xu F, Fu J, Li S: Protective roles of selenium on nitric oxide and the gene expression of inflammatory cytokines induced by cadmium in chicken splenic lymphocytes. Biol Trace Elem Res 2015;168:252-260. https://doi.org/10.1007/s12011-015-0354-z |
||||
| 11 Du Q, Yao H, Yao L, Zhang Z, Lei X, Xu S: Selenium deficiency influences the expression of selenoproteins and inflammatory cytokines in chicken aorta vessels. Biol Trace Elem Res 2016;173:501-513. https://doi.org/10.1007/s12011-016-0676-5 |
||||
| 12 Yang CC, Yao CA, Lin YR, Yang JC, Chien CT: Deep-sea water containing selenium provides intestinal protection against duodenal ulcers through the upregulation of Bcl-2 and thioredoxin reductase 1. PLoS One 2014;9:e96006. https://doi.org/10.1371/journal.pone.0096006 |
||||
| 13 Liu ZW, Zhu HT, Chen KL, Qiu C, Tang KF, Niu XL: Selenium attenuates high glucose-induced ROS/TLR-4 involved apoptosis of rat cardiomyocyte. Biol Trace Elem Res 2013;156:262-270. https://doi.org/10.1007/s12011-013-9857-7 |
||||
| 14 Pei Z, Li H, Guo Y, Jin Y, Lin D: Sodium selenite inhibits the expression of VEGF, TGFβ1 and IL-6 induced by LPS in human PC3 cells via TLR4-NF-κB signaling blockage. Int Immunopharmacol 2010;10:50-56. https://doi.org/10.1016/j.intimp.2009.09.020 |
||||
| 15 Kudva AK, Shay AE, Prabhu KS: Selenium and inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol 2015;309:G71-G77. https://doi.org/10.1152/ajpgi.00379.2014 |
||||
| 16 Castro Aguilar-Tablada T, Navarro-Alarcón M, Quesada Granados J, Samaniego Sánchez C, Rufián-Henares J, Nogueras-Lopez F: Ulcerative colitis and Crohn's disease are associated with decreased serum selenium concentrations and increased cardiovascular risk. Nutrients 2016;8:780. https://doi.org/10.3390/nu8120780 |
||||
| 17 Han YM, Yoon H, Lim S, Sung MK, Shin CM, Park YS, Kim N, Lee DH, Kim JS: Risk factors for vitamin D, zinc, and selenium deficiencies in Korean patients with inflammatory bowel disease. Gut Liver 2017;11:363. https://doi.org/10.5009/gnl16333 |
||||
| 18 Barrett CW, Singh K, Motley AK, Lintel MK, Matafonova E, Bradley AM, Ning W, Poindexter SV, Parang B, Reddy VK: Dietary selenium deficiency exacerbates DSS-induced epithelial injury and AOM/DSS-induced tumorigenesis. PLoS One 2013;8:e67845. https://doi.org/10.1371/journal.pone.0067845 |
||||
| 19 Sang L, Chang B, Zhu J, Yang F, Li Y, Jiang X, Sun X, Lu C, Wang D: Dextran sulfate sodium-induced acute experimental colitis in C57BL/6 mice is mitigated by selenium. Int Immunopharmacol 2016;39:359-368. https://doi.org/10.1016/j.intimp.2016.07.034 |
||||
| 20 Bitiren M, Karakilcik AZ, Zerin M, Ozardalı I, Selek S, Nazlıgül Y, Ozgonul A, Musa D, Uzunkoy A: Protective effects of selenium and vitamin E combination on experimental colitis in blood plasma and colon of rats. Biol Trace Elem Res 2010;136:87-95. https://doi.org/10.1007/s12011-009-8518-3 |
||||
| 21 Ala M, Kheyri Z: The rationale for selenium supplementation in inflammatory bowel disease: A mechanism-based point of view. Nutrition 2021:111153. https://doi.org/10.1016/j.nut.2021.111153 |
||||
| 22 Keshavarzian A, Haydek J, Zabihi R, Doria M, D'Astice M, Sorenson J: Agents capable of eliminating reactive oxygen species. Catalase, WR-2721, or Cu (II) 2 (3,5-DIPS) 4 decrease experimental colitis. Dig Dis Sci 1992;37:1866-1873. https://doi.org/10.1007/BF01308081 |
||||
| 23 Jafari RM, Ghahremani MH, Rahimi N, Shadboorestan A, Rashidian A, Esmaeili J, Mehr SE, Dehpour AR: The anticonvulsant activity and cerebral protection of chronic lithium chloride via NMDA receptor/nitric oxide and phospho-ERK. Brain Res Bull 2018;137:1-9. https://doi.org/10.1016/j.brainresbull.2017.10.015 |
||||
| 24 Duntas L: Selenium and inflammation: underlying anti-inflammatory mechanisms. Horm Metab Res 2009;41:443-447. https://doi.org/10.1055/s-0029-1220724 |
||||
| 25 Bi CL, Wang H, Wang YJ, Sun J, Dong JS, Meng X, Li JJ: Selenium inhibits Staphylococcus aureus-induced inflammation by suppressing the activation of the NF-κB and MAPK signalling pathways in RAW264.7 macrophages. Eur J Pharmacol 2016;780:159-165. https://doi.org/10.1016/j.ejphar.2016.03.044 |
||||
| 26 Liu T, Zhang L, Joo D, Sun SC: NF-κB signaling in inflammation. Signal Transduct Target Ther 2017;2:17023. https://doi.org/10.1038/sigtrans.2017.23 |
||||
| 27 Abraham C, Medzhitov R: Interactions between the host innate immune system and microbes in inflammatory bowel disease. Gastroenterology 2011;140:1729-1737. https://doi.org/10.1053/j.gastro.2011.02.012 |
||||
| 28 Abraham C, Cho J: Interleukin-23/Th17 pathways and inflammatory bowel disease. Inflamm Bowel Dis 2009;15:1090-1100. https://doi.org/10.1002/ibd.20894 |
||||
| 29 Karban AS, Okazaki T, Panhuysen CI, Gallegos T, Potter JJ, Bailey-Wilson JE, Silverberg MS, Duerr RH, Cho JH, Gregersen PK: Functional annotation of a novel NFKB1 promoter polymorphism that increases risk for ulcerative colitis. Hum Mol Genet 2004;13:35-45. https://doi.org/10.1093/hmg/ddh008 |
||||
| 30 Glas J, Török HP, Tonenchi L, Müller-Myhsok B, Mussack T, Wetzke M, Klein W, Epplen JT, Griga T, Schiemann U: Role of the NFKB1 −94ins/delATTG promoter polymorphism in IBD and potential interactions with polymorphisms in the CARD15/NOD2, IKBL, and IL-1RN genes. Inflamm Bowel Dis 2006;12:606-611. https://doi.org/10.1097/01.ibd.0000225346.23765.6b |
||||
| 31 Gao X, Zhang Z, Xing H, Yu J, Zhang N, Xu S: Selenium deficiency-induced inflammation and increased expression of regulating inflammatory cytokines in the chicken gastrointestinal tract. Biol Trace Elem Res 2016;173:210-218. https://doi.org/10.1007/s12011-016-0651-1 |
||||
| 32 Jena G, Trivedi PP, Sandala B: Oxidative stress in ulcerative colitis: an old concept but a new concern. Free Radic Res 2012;46:1339-1345. https://doi.org/10.3109/10715762.2012.717692 |
||||
| 33 Piechota-Polanczyk A, Fichna J: Review article: the role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn-Schmiedeberg Arch Pharmacol 2014;387:605-620. https://doi.org/10.1007/s00210-014-0985-1 |
||||
| 34 Hassan A, Ibrahim A, Mbodji K, Coëffier M, Ziegler F, Bounoure F, Chardigny JM, Skiba M, Savoye G, Déchelotte P: An α-linolenic acid-rich formula reduces oxidative stress and inflammation by regulating NF-κB in rats with TNBS-induced colitis. J Nutr 2010;140:1714-1721. https://doi.org/10.3945/jn.109.119768 |
||||
| 35 Chen S, Chen H, Du Q, Shen J: Targeting myeloperoxidase (MPO) mediated oxidative stress and inflammation for reducing brain ischemia injury: Potential application of natural compounds. Front Physiol 2020;11:433. https://doi.org/10.3389/fphys.2020.00433 |
||||
| 36 Zhu H, Li YR: Oxidative stress and redox signaling mechanisms of inflammatory bowel disease: updated experimental and clinical evidence. Exp Biol Med 2012;237:474-480. https://doi.org/10.1258/ebm.2011.011358 |
||||
| 37 Han X, Lee A, Huang S, Gao J, Spence JR, Owyang C: Lactobacillus rhamnosus GG prevents epithelial barrier dysfunction induced by interferon-gamma and fecal supernatants from irritable bowel syndrome patients in human intestinal enteroids and colonoids. Gut Microbes 2019;10:59-76. https://doi.org/10.1080/19490976.2018.1479625 |
||||
| 38 Lee SH: Intestinal permeability regulation by tight junction: implication on inflammatory bowel diseases. Intest Res 2015;13:11. https://doi.org/10.5217/ir.2015.13.1.11 |
||||
| 39 Fretland D, Widomski D, Anglin C, Walsh R, Levin S, Riley D, Weiss R, Gaginella T: Superoxide dismutase (SOD) modulates acetic acid-induced colitis in rodents. Gastroenterology 1991;100:A581. https://doi.org/10.1016/0891-5849(90)90680-H |
||||
| 40 Shaker ME, Ashamallah SA, Houssen ME: Celastrol ameliorates murine colitis via modulating oxidative stress, inflammatory cytokines and intestinal homeostasis. Chem Biol Interact 2014;210:26-33. https://doi.org/10.1016/j.cbi.2013.12.007 |
||||
| 41 Zhou Z, Dai C, Liu W: Anti-TNF-A therapy about infliximab and adalimamab for the effectiveness in ulcerative colitis compared with conventional therapy: a meta-analysis. Hepatogastroenterology 2015;62:309-318. | ||||
| 42 Chen B, Alvarado DM, Iticovici M, Kau NS, Park H, Parikh PJ, Thotala D, Ciorba MA: Interferon-induced IDO1 mediates radiation resistance and is a therapeutic target in colorectal cancer. Cancer Immunol Res 2020;8:451-464. https://doi.org/10.1158/2326-6066.CIR-19-0282 |
||||
| 43 Hosseini-Tabatabaei A, Jalili RB, Li Y, Kilani RT, Rezakhanlou AM, Ghahary A: Mechanism underlying defective interferon gamma-induced IDO expression in non-obese diabetic mouse fibroblasts. PLoS One 2012;7:e37747. https://doi.org/10.1371/journal.pone.0037747 |
||||
| 44 Ala M: Tryptophan metabolites modulate inflammatory bowel disease and colorectal cancer by affecting immune system. Int Rev Immunol 2021:1-20. https://doi.org/10.1080/08830185.2021.1954638 |
||||
| 45 Sofia MA, Ciorba MA, Meckel K, Lim CK, Guillemin GJ, Weber CR, Bissonnette M, Pekow JR: Tryptophan metabolism through the kynurenine pathway is associated with endoscopic inflammation in ulcerative colitis. Inflamm Bowel Dis 2018;24:1471-1480. https://doi.org/10.1093/ibd/izy103 |
||||
| 46 Gupta NK, Thaker AI, Kanuri N, Riehl TE, Rowley CW, Stenson WF, Ciorba MA: Serum analysis of tryptophan catabolism pathway: correlation with Crohn's disease activity. Inflamm Bowel Dis 2012;18:1214-1220. https://doi.org/10.1002/ibd.21849 |
||||
| 47 Lamas B, Natividad JM, Sokol H: Aryl hydrocarbon receptor and intestinal immunity. Mucosal Immunol 2018;11:1024-1038. https://doi.org/10.1038/s41385-018-0019-2 |
||||
| 48 Gao J, Xu K, Liu H, Liu G, Bai M, Peng C, Li T, Yin Y: Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Front Cell Infect Microbiol 2018;8:13. https://doi.org/10.3389/fcimb.2018.00013 |
||||
| 49 Ouyang W, Valdez P: IL-22 in mucosal immunity. Mucosal Immunol 2008;1:335-338. https://doi.org/10.1038/mi.2008.26 |
||||
| 50 Yamamoto-Furusho JK, Miranda-Pérez E, Fonseca-Camarillo G, Sánchez-Muñoz F, Dominguez-Lopez A, Barreto-Zuñiga R: Colonic epithelial upregulation of interleukin 22 (IL-22) in patients with ulcerative colitis. Inflamm Bowel Dis 2010;16:1823-1823. https://doi.org/10.1002/ibd.21235 |
||||
| 51 Schmechel S, Konrad A, Diegelmann J, Glas J, Wetzke M, Paschos E, Lohse P, Göke B, Brand S: Linking genetic susceptibility to Crohn's disease with Th17 cell function: IL-22 serum levels are increased in Crohn's disease and correlate with disease activity and IL23R genotype status. Inflamm Bowel Dis 2008;14:204-212. https://doi.org/10.1002/ibd.20315 |
||||
| 52 Brand S, Beigel F, Olszak T, Zitzmann K, Eichhorst ST, Otte JM, Diepolder H, Marquardt A, Jagla W, Popp A, Leclair S, Herrmann K, Seiderer J, Ochsenkühn T, Göle B, Auernhammer CJ, Dambacher J: IL-22 is increased in active Crohn's disease and promotes proinflammatory gene expression and intestinal epithelial cell migration. Am J Physiol Gastrointest Liver Physiol 2006;290:G827-G838. https://doi.org/10.1152/ajpgi.00513.2005 |
||||
| 53 Nikolaus S, Schulte B, Al-Massad N, Thieme F, Schulte DM, Bethge J, Rehman A, Tran F, Aden K, Häsler R: Increased tryptophan metabolism is associated with activity of inflammatory bowel diseases. Gastroenterology 2017;153:1504-1516.e2. https://doi.org/10.1053/j.gastro.2017.08.028 |
||||
| 54 Feng T, Qin H, Wang L, Benveniste EN, Elson CO, Cong Y: Th17 cells induce colitis and promote Th1 cell responses through IL-17 induction of innate IL-12 and IL-23 production. J Immunol 2011;186:6313-6318. https://doi.org/10.4049/jimmunol.1001454 |
||||
| 55 Fukata M, Shang L, Santaolalla R, Sotolongo J, Pastorini C, España C, Ungaro R, Harpaz N, Cooper HS, Elson G: Constitutive activation of epithelial TLR4 augments inflammatory responses to mucosal injury and drives colitis-associated tumorigenesis. Inflamm Bowel Dis 2010;17:1464-1473. https://doi.org/10.1002/ibd.21527 |
||||
| 56 Rashidian A, Muhammadnejad A, Dehpour AR, Mehr SE, Akhavan MM, Shirkoohi R, Chamanara M, Mousavi SE, Rezayat SM: Atorvastatin attenuates TNBS-induced rat colitis: the involvement of the TLR4/NF-kB signaling pathway. Inflammopharmacology 2016;24:109-118. https://doi.org/10.1007/s10787-016-0263-6 |
||||
| 57 Chamanara M, Rashidian A, Mehr SE, Dehpour AR, Shirkohi R, Akbarian R, Abdollahi A, Rezayat SM: Melatonin ameliorates TNBS-induced colitis in rats through the melatonin receptors: involvement of TLR4/MyD88/NF-κB signalling pathway. Inflammopharmacology 2019;27:361-371. https://doi.org/10.1007/s10787-018-0523-8 |
||||
| 58 Zhang J, Dou W, Zhang E, Sun A, Ding L, Wei X, Chou G, Mani S, Wang Z: Paeoniflorin abrogates DSS-induced colitis via a TLR4-dependent pathway. Am J Physiol Gastrointest Liver Physiol 2013;306:G27-G36. https://doi.org/10.1152/ajpgi.00465.2012 |
||||
| 59 Gupta RA, Motiwala MN, Dumore NG, Danao KR, Ganjare AB: Effect of piperine on inhibition of FFA induced TLR4 mediated inflammation and amelioration of acetic acid induced ulcerative colitis in mice. J Ethnopharmacol 2015;164:239-246. https://doi.org/10.1016/j.jep.2015.01.039 |
||||
| 60 He X, Wei Z, Wang J, Kou J, Liu W, Fu Y, Yang Z: Alpinetin attenuates inflammatory responses by suppressing TLR4 and NLRP3 signaling pathways in DSS-induced acute colitis. Sci Rep 2016;6:28370. https://doi.org/10.1038/srep28370 |
||||
| 61 Kang GD, Kim DH: Poncirin and its metabolite ponciretin attenuate colitis in mice by inhibiting LPS binding on TLR4 of macrophages and correcting Th17/Treg imbalance. J Ethnopharmacol 2016;189:175-185. https://doi.org/10.1016/j.jep.2016.05.044 |
||||
| 62 Sun J, Zhang H, Guan L, Zhou H, Sun M: Alpha-lipoic acid attenuates trinitrobenzene sulfonic acid-induced ulcerative colitis in mice. Int J Clin Exp Med 2015;8:358. | ||||
| 63 Qin L, Yao ZQ, Chang Q, Zhao YL, Liu NN, Zhu XS, Liu QQ, Wang LF, Yang AG, Gao CF: Swimming attenuates inflammation, oxidative stress, and apoptosis in a rat model of dextran sulfate sodium-induced chronic colitis. Oncotarget 2017;8:7391. https://doi.org/10.18632/oncotarget.14080 |
||||
| 64 Rahman MM, Uson-Lopez RA, Sikder MT, Tan G, Hosokawa T, Saito T, Kurasaki M: Ameliorative effects of selenium on arsenic-induced cytotoxicity in PC12 cells via modulating autophagy/apoptosis. Chemosphere 2018;196:453-466. https://doi.org/10.1016/j.chemosphere.2017.12.149 |
||||
| 65 El Asar HM, Mohammed EA, Aboulhoda BE, Emam HY, Imam AAA: Selenium protection against mercury neurotoxicity: Modulation of apoptosis and autophagy in the anterior pituitary. Life Sci 2019:116578. https://doi.org/10.1016/j.lfs.2019.116578 |
||||