×
![]()
Corresponding Author: Salvatore Cuzzocrea and Rosanna Di Paola
Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Viale Ferdinando Stagno D’Alcontres 31, 98166 Messina (Italy)
E-Mail salvator@unime.it
Department of Veterinary Sciences, University of Messina, Polo Universitario dell’Annunziata, 98168 Messina (Italy)
E-Mail dipaolar@unime.it
Açaí (Euterpe Oleraceae Mart.) Seeds Regulate NF-κB and Nrf2/ARE Pathways Protecting Lung against Acute and Chronic Inflammation
Tiziana Genovesea Ramona D’Amicoa Roberta Fuscob Daniela Impellizzeria Alessio Filippo Peritorea Rosalia Crupic Livia Interdonatoa Enrico Gugliandoloc Salvatore Cuzzocreaa,d Rosanna Di Paolac Rosalba Siracusaa Marika Cordaroe
aDepartment of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Messina, Italy, bDepartment of Clinical and Experimental Medicine, University of Messina, Messina, Italy, cDepartment of Veterinary Sciences, University of Messina, Polo Universitario dell’Annunziata, Messina, Italy, dDepartment of Pharmacological and Physiological Science, Saint Louis University School of Medicine, Saint Louis, MO, USA, eDepartment of Biomedical, Dental and Morphological and Functional Imaging, University of Messina, Messina, Italy
Introduction
Acute and chronic lung diseases are caused by inflammation in the lungs. Acute lung injury (ALI) is defined by a fast inflammatory response in the lungs caused by either communicable (infection) or non-communicable (acid aspiration) disorders [1]. As the lung is exposed to external factors either through the airways such as infectious agents and/or environmental pollutants or through the blood stream like sepsis and/or endotoxin, ALI can result in acute respiratory distress syndrome (ARDS), a spectrum of lung diseases characterized by a severe inflammatory process in the lung parenchyma causing diffuse alveolar damage and respiratory failure [2-4]. When acute inflammation does not resolve completely, chronic inflammation develops. Chronic inflammatory responses clean necrotic debris and apoptotic cells from acute inflammation, guard against and inhibit infection spread, and heal and repair lung tissue damage [5]. Pneumonia and acute respiratory distress syndrome (ARDS) are examples of acute inflammation, whereas asthma and chronic obstructive lung disease (COPD) are examples of chronic inflammation [6].
Most airway pathologies have in common an implementation of inflammatory pathways activation and an increase oxidative stress in various kinds of respiratory cells. Intracellular buildup of reactive oxygen species (ROS) can damage macromolecules such as DNA, proteins, and lipids, resulting in cell abnormalities or death, on the other hands, inflammatory reactions are triggered by unbalanced redox equilibrium and cell damage, which exacerbate oxidative stress [7]. One of the most regulators that protects cells from ROS and xenobiotics is NF-E2-related factor 2 (Nrf2), a cap ‘n’ collar (CNC)-bZIP family transcription factor. Nrf2 binds to antioxidant response elements (AREs) and activates a battery of antioxidant and detoxifying genes when activated by oxidative and xenobiotic stressors [8]. Nrf2 also activates several developmental genes, allowing it to control cell proliferation, differentiation, cell death, and metabolic programs [9, 10]. Furthermore, Nrf2 interacts with a variety of other proteins and pathways, forming a complex regulatory network for redox homeostasis and other cellular functions. For a variety of human disorders, including respiratory ailments, Nrf2 has been identified as a crucial factor and an important therapeutic target [11]. Additionally, under oxidative stress, complex biological pathways (transcriptional and post-transcriptional) connect Nrf2 and NF-κB. Nrf2 deficiency increase NF-κB activity, which promotes cytokine release as well as an increase in molecules adhesion production and more [12].
For these reasons the development of Nrf2-NF-κB targeting therapeutics will be greatly aided by a thorough knowledge of the molecular partners and downstream activities of Nrf2.
Glucocorticosteroids are often used as an anti-inflammatory therapy for acute and chronic lung inflammation, but specific nutrients have been proposed to positively affect disease progression as novel therapy alternatives for individuals suffering from major side effects or developing resistance [13-15].
In the last years, the foods that has been attracting the attention of researchers in recent years is Acai seeds. Acai Berry is a berry with various nutritional properties and a certain therapeutic potential. This fruit, with a sour and pleasant taste, is produced from a palm (Euterpe Oleracea), widespread only in the Amazon. Considered a highly energetic food, the Acai fruit has been consumed by the Indians of the Amazon for centuries, as a food base and natural remedy for various diseases.
Recently, for its antioxidant, energizing and potentially slimming properties, Acai Berry has gained popularity as a functional ingredient, especially following the marketing of its derivatives in the form of juices and food supplements. Currently, the known effects of Acai are mainly determined by the high content of antioxidants, unsaturated fats (omega-3, omega-6 and omega-9), vitamins, minerals such as iron and calcium, that make it a fruit hypocholesterolemic, immunostimulant, anti-inflammatory and antioxidant [16-24].
However, from a scientific point of view, the evidence supporting the positive health effects is still scarce. The research is oriented towards the study of the other possible properties of the Açai Seeds, in particular on its antioxidant activity and on its potential effects in the prevention of diseases caused by oxidative stress. Procyanidins (dimers up to pentamers), protocatechuic acid, and epicatechin were the most prevalent molecules found in previous investigations and identification of chemicals in Açai Seed extracts. Furthermore, seed extracts had comparable or better scavenging ability against peroxyl, peroxynitrites, and hydroxyl radicals than the pulp, which they linked to the observed procyanidins [19, 25, 26].
With these background in our mind, in this work we tested for the first time the beneficial effects of Açai Seeds assumption in an acute and chronic model of lung inflammation.
Materials and Methods
Animals
CD1 mice (25-30 g, Envigo, Milan, Italy) were employed. The University of Messina Review Board for animal care (OPBA) approved the study (Ethical Code: 89126.24/2021-PR). All animal experiments agree with the new Italian regulations (D.Lgs 2014/26), EU regulations (EU Directive 2010/63) and the ARRIVE guidelines.
Experimental Design and Groups
First experimental design. ALI was made as previously described by Di Paola et al. [27]. Briefly, mice were anaesthetized and saline (0.1 mL) or saline containing 2% λ carrageenan (CAR) (0.1 ml) was injected into the pleural cavity. Açai seeds were washed with water and frozen at −80 °C. The frozen seed was lyophilized and stored at −20 °C as previously described [28]. Powder obtained were following dissolved in saline.
Mice were casually distributed into the following groups:
(1) CAR: mice were subjected to the CAR injection described above, and treated with saline solution;
(2) CAR+Açai Seeds: mice were subjected to the CAR injection described above, and treated orally 1 h after with Açai Seeds dissolved in saline at different doses (100 mg/kg, 300 mg/kg and 500 mg/kg);
(3) Sham group: animals were subjected to an injection of saline solution;
(4) Sham groups+Açai Seeds: animals were treated with Açai Seeds dissolved in saline at different doses (100 mg/kg, 300 mg/kg and 500 mg/kg) (data not shown).
In another set of experiments, we compare the highest dose of Açai Seeds with Resveratrol dissolved in saline and administered by oral route at the dose of 50 mg/kg (Supplementary Fig. S1 – for all supplementary material see www.cellphysiolbiochem.com) [29].
After four hours, animals were killed by cervical dislocation under anesthesia and lungs were conserved for further study. Doses were chosen based on a dose-response study carried out in our lab. Açai Seeds at the doses of 100 mg/kg and 300 mg/kg did not show any advantage in CAR-induced lung inflammation. For this reason, we carried on our experiment only with the dose of 500 mg/kg.
Second experimental design. Bleomycin administration was performed as previously described to reproduce lung fibrosis [30, 31]. In detail, bleomycin sulphate (1mg/kg body weight) was delivered by a single intratracheal administration. A volume of 100 mL was injected at end-expiration to guarantee delivery to the distal airways. This was immediately followed by 300 mL of air [32-34].
Mice were randomly divided into groups:
(I) Bleomycin: animals that receive one injection of bleomycin.
(IV) Bleomycin+ Açai Seeds: mice were subjected to the bleomycin injection described above and treated orally with Açai Seeds dissolved in saline at the dose of 500 mg/kg for 14 days.
(I) Sham: animals that were exposed to the vehicle (saline with 10% of DMSO).
At the end of experiment, mice were sacrificed by cervical dislocation under anesthesia, and lung tissue and bronchoalveolar lavage fluid (BALF) were collected as previously described [35-40].
Exudate and leukocytes count
Measure of exudates and count of leukocytes was made as previously described [41]. Briefly, at the end of the experiment, the chest was opened, and the pleural cavity washed with 1 mL of saline solution containing heparin and indomethacin. The exudate and washing solution were removed by aspiration and the total volume measured. The leukocytes in the exudate were suspended in phosphate-buffer saline (PBS) and counted with after Blue Toluidine staining an optical microscope in a Burker’s chamber.
Measurement of Lung Edema
At the end of the experiment, wet lung weights were recorded. The lungs were subsequently dried for 48 h at 180 °C and then weighed again. The water content of the lungs was calculated as the ratio of the wet:dry weight of the tissue [29-31, 42].
Bronchoalveolar Lavage (BAL)
At the end of experiment, mice were euthanized, and the tracheas were cannulated to perform the lavage for cell counting, as previously described [29-31, 42]. Additionally, from BAL, we analyzed the total protein content using a DC Protein Assay kit (Bio-Rad Laboratories, Hercules, CA, USA) as previously described [43, 44].
Western Blot Analysis of Cytosolic and Nuclear Extracts
Extracts of the cytosol and nucleus were prepared, as previously mentioned [45-49]. The following primary antibodies were used: anti-iNOS (1:500, Santa Cruz Biotechnology, #sc-7271), anti-NRF-2 (1-500, Santa Cruz Biotechnology, Heidelberg, Germany, #sc-365949), anti-heme oxygenase 1 (HO-1; 1-500, Santa Cruz Biotechnology, Heidelberg, Germany, #sc-136960), anti-(α-sma antibody (1:250, Santa Cruz Biotechnology), anti-Iκbα (1:500, Santa Cruz Biotechnology, #sc-1643), and anti-nfκb (1:500, Santa Cruz Biotechnology, #sc8414) in 1× PBS, 5% w/v non-fat dried milk, and 0.1% Tween 20, at 4 °C overnight [42, 50-52]. For the cytosolic fraction, Western blots were also explored with antibody against β-actin protein (1:500, Santa Cruz Biotechnology, Dallas, TX, USA). The same methods were used for nuclear fraction with lamin A/C (1:500, Sigma-Aldrich Corp., Milan, Italy) [53, 54]. Signals were examined with an enhanced chemiluminescence (ECL) detection system reagent, according to the manufacturer’s instructions (Thermo, Monza, Italy). The relative expression of the protein bands was quantified by densitometry with BIORAD ChemiDocTM XRS+ software [46, 50, 55-57].
Evaluation of Tissue Lipid Peroxidation
Lipid peroxidation were assessed with the consolidated method of malonaldehyde as previously described and expressed as mM MDA/mg of proteins [58].
Cytokine and Nitrite/Nitrate Measurement
TNF-a or IL-6 or IL-1b and nitrite/nitrate concentration were measured in the exudates using ELISA kits (R&D Systems, Minneapolis, MN, USA) following the manufacturer’s instructions and expressed as pg/mL or with Griess reaction and express as mmol/mouse [27, 55, 56, 58-64].
Assessment of antioxidant system
The SOD activity assay was measured following the method described by Marklund and Marklund at 420 nm [65]. The results are presented as SOD units/mg protein. Moreover, the activity of CAT was assessed following the method of Aebi in terms of mmoles of hydrogen peroxide (H2O2) consumed per min per mg of protein [66]. The results are presented as CAT units/mg protein. Furthermore, the level of the nonenzymatic cellular antioxidant glutathione was measured following the method of Moron et al. [67-69]. The results are presented as GSH units/mg protein.
Measurement of Reactive Oxygen Species (ROS)
2’,7’-dichlorofluorescein diacetate (Molecular Probes, Eugene, OR, USA) was used to detect ROS production in lung tissue homogenates as previously described [70-72]. The results were normalized as ROS produced per milligram of protein.
Histopathological Evaluation with Hematoxylin/Eosin and Toluidine Blue and masson
The lung tissues were stained with hematoxylin and eosin (H&E) for architecture alterations or toluidine blue for mast cells degranulation, or with Masson Thrichrome for collagen deposition and analyzed using a light microscopy (Leica DM6, Milan, Italy) associated with an Imaging system (LasX Navigator, Milan, Italy) [46, 49, 50, 73].
Immunohistochemical localization of Nitrotyrosine, poly(ADP-ribose), ICAM and P-Selectin
At the end of experiments lung tissue were incubated with anti-ICAM-1 murine polyclonal antibody (1/100 in PBS, v/v, Santa Cruz Biotechnology), anti-P-selectin murine polyclonal antibody (1/100 in PBS, v/v, Santa Cruz Biotechnology), anti-PAR murine polyclonal antibody (1/100 in PBS, v/v, Santa Cruz Biotechnology) and anti-nitrotyrosine rabbit polyclonal antibody (1:200 in PBS, v/v, Millipore) as previously described [29-31, 42, 53, 57, 74-77]. Immunohistochemical images were collected using (Leica DM6, Milan, Italy) associated with an Imaging system (LasX Navigator, Milan, Italy). The digital images were opened in ImageJ, followed by deconvolution using the color deconvolution plug-in. When the IHC profiler plug-in is selected, it automatically plots a histogram profile of the deconvoluted DAB image, and a corresponding scoring log is displayed. The histogram profile corresponds to the positive pixel intensity value obtained from the computer program. All immunohistochemical analyses were carried out by two observers blinded to the treatment [29-31, 42, 74-78].
Materials
Unless otherwise stated, all compounds were purchased from Sigma-Aldrich.
Statistical Evaluation
In this study, the data are expressed as the average ± SEM and represent at least 3 experiments carried out in different days. For in vivo studies, N represents the number of animals used. The number of animals used for in vivo studies was carried out by G*Power 3.1 software (Die Heinrich-Heine-Universität Düsseldorf, Düsseldorf, Germany). Data were analyzed by an experienced histopathologist, and all the studies were performed without knowledge of the treatments. The results were analyzed by one-way ANOVA followed by a Bonferroni post-hoc test for multiple comparisons. A p value less than 0.05 was considered significant.
Results
Açai Seeds administration reduce histological damage CAR-induced
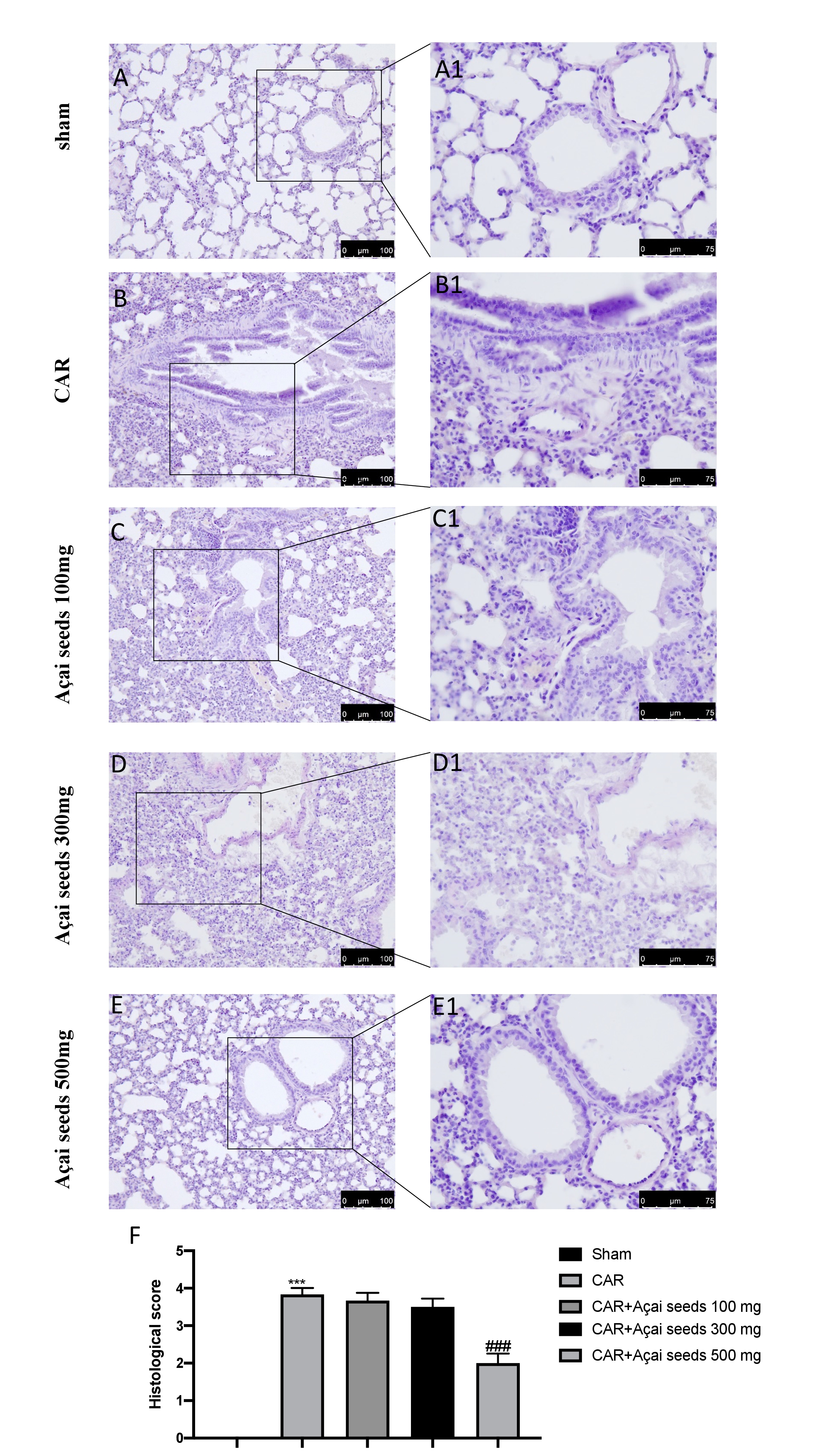
Architecture alteration, infiltration of neutrophils and oedema are the main characteristic of ALI as showed in Fig. 1 (B-B1 see histological score 1F) compared to sham animals (Fig. 1A-A1 see histological score 1F). Açai Seeds at the doses of 100 mg/Kg (Fig. 1C-C1 see histological score 1F) and 300 mg/Kg (Fig. 1D-D1 see histological score 1F) not showed any significantly decrease in the inflammatory state. On the other hand, the higher dose used, that is 500 mg/kg (Fig. 1E-E1 see histological score 1G), showed a significantly reduction in the degree of injury CAR-induced.
Açai Seeds administration reduce exudates and cytokines release
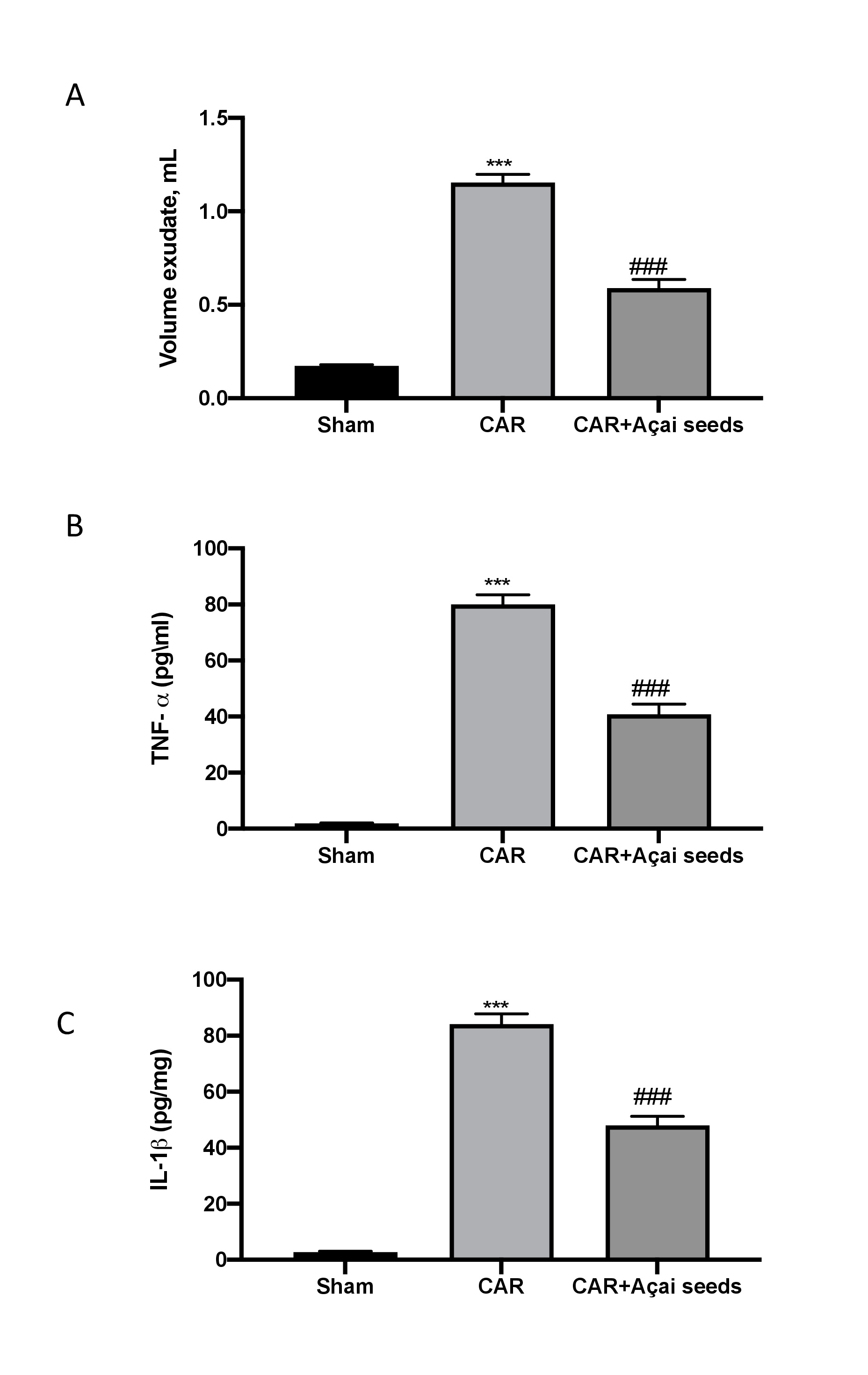
Injection of CAR stimulated an acute inflammatory response characterized by the accumulation of exudates (Fig. 2A) rich in content of pro-inflammatory cytokines TNF-a (Fig. 2B) as well as IL-1b (Fig. 2C). Açai Seeds administered orally at the dose of 500 mg/kg was able to significantly reduced exudates formation as well as pro-inflammatory cytokines production.
Açai Seeds inhibits mast cells degranulation, PMNs infiltration and MPO production
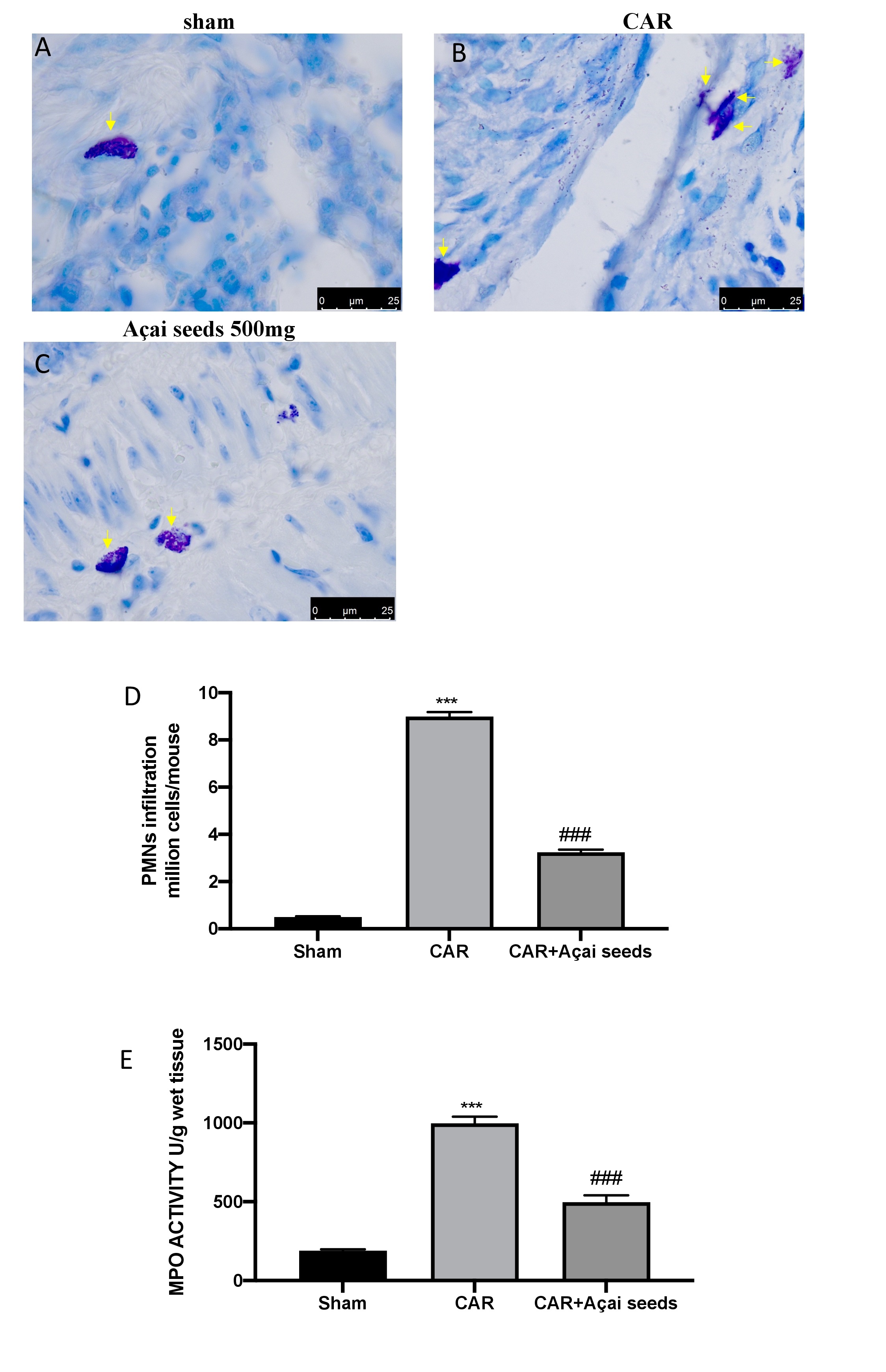
ALI was also characterized by a significant influx of mast cells, PMNs, and leukocytes into the interstitium and bronchoalveolar space. As believed, after CAR injection we found a significantly increase in mast cell degranulation, PMNs infiltration and MPO levels (Fig. 3B, 3D and 3E) compared to sham animals (Fig. 3A, 3D and 3E) [2]. On the other way the administration of Açai Seeds at the dose of 500 mg/kg significantly decreases all these parameters.
Açai Seeds administration decrease inflammation and improve antioxidant defense
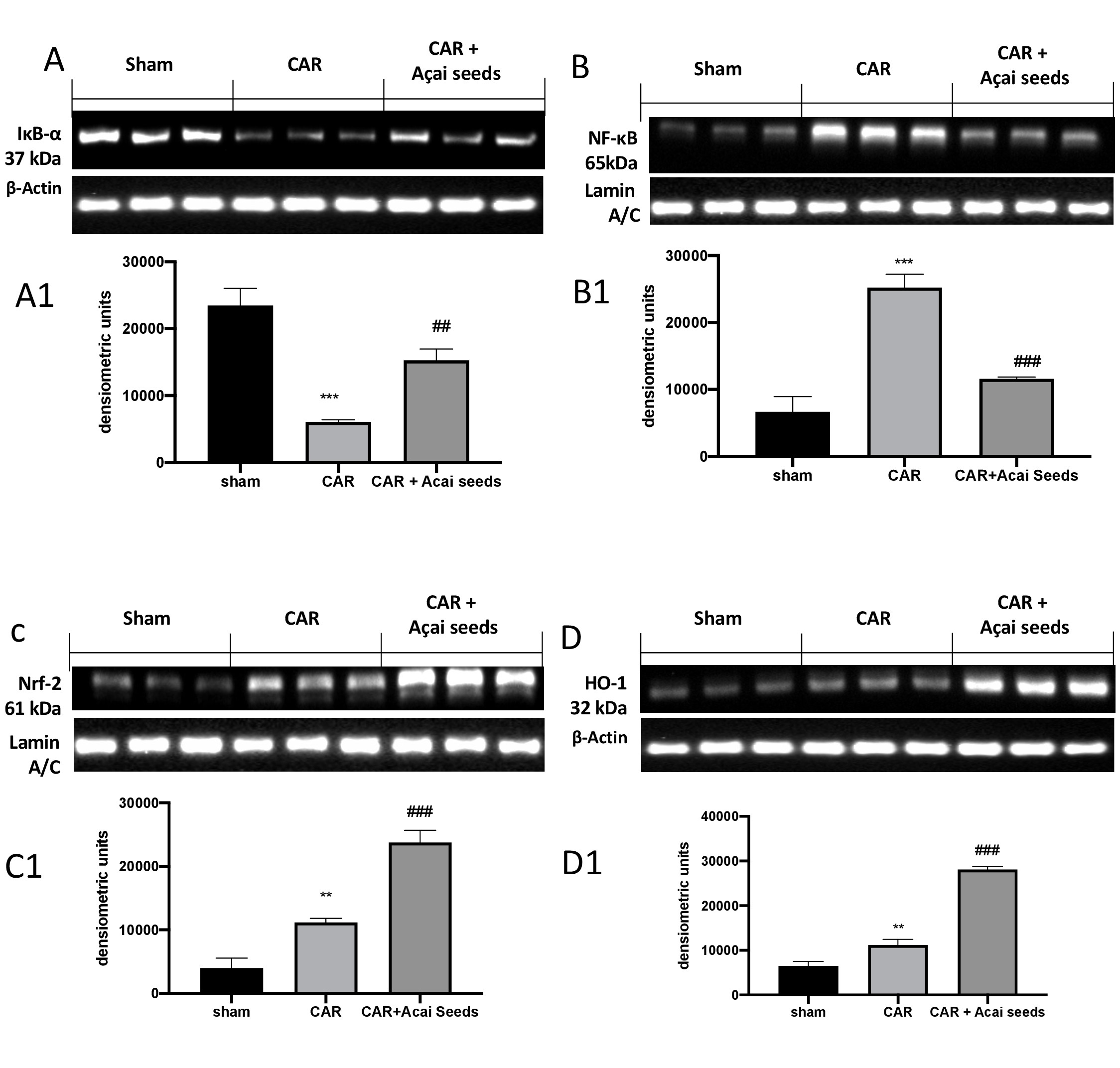
To understand a possible molecular pathway by which Açai Seeds act, we investigated by western blots the activation of NF-κB pathways. As showed in Fig. 4, we found that following CAR-injection there were a significantly decrease in IκB-a (Fig. 4A and relative densitometric analysis A1) and therefore a significantly increase in NF-κB expression compared to sham group (Fig. 4B and relative densitometric analysis B1). Açai Seeds administration at the dose of 500 mg/kg was able to restore in IκB-a (Fig. 4A and relative densitometric analysis A1) expression as well as to decrease NF-κB translocation (Fig. 4B and relative densitometric analysis B1). Additionally, considering to anti-oxidative property of Açai Seeds, we investigated Nrf-2 pathways, always by western blot. We found that following CAR-injection into pleural cavity, there were a small but significant increase in Nrf-2 expression (Fig. 4C and relative densitometric analysis C1) as well as in HO-1 expression (Fig. 4 D and relative densitometric analysis D1). After, Açai Seeds administration both expression of Nrf-2 (Fig. 4C and relative densitometric analysis C1) and HO-1 expression (Fig. 4D and relative densitometric analysis D1) were significantly improved.
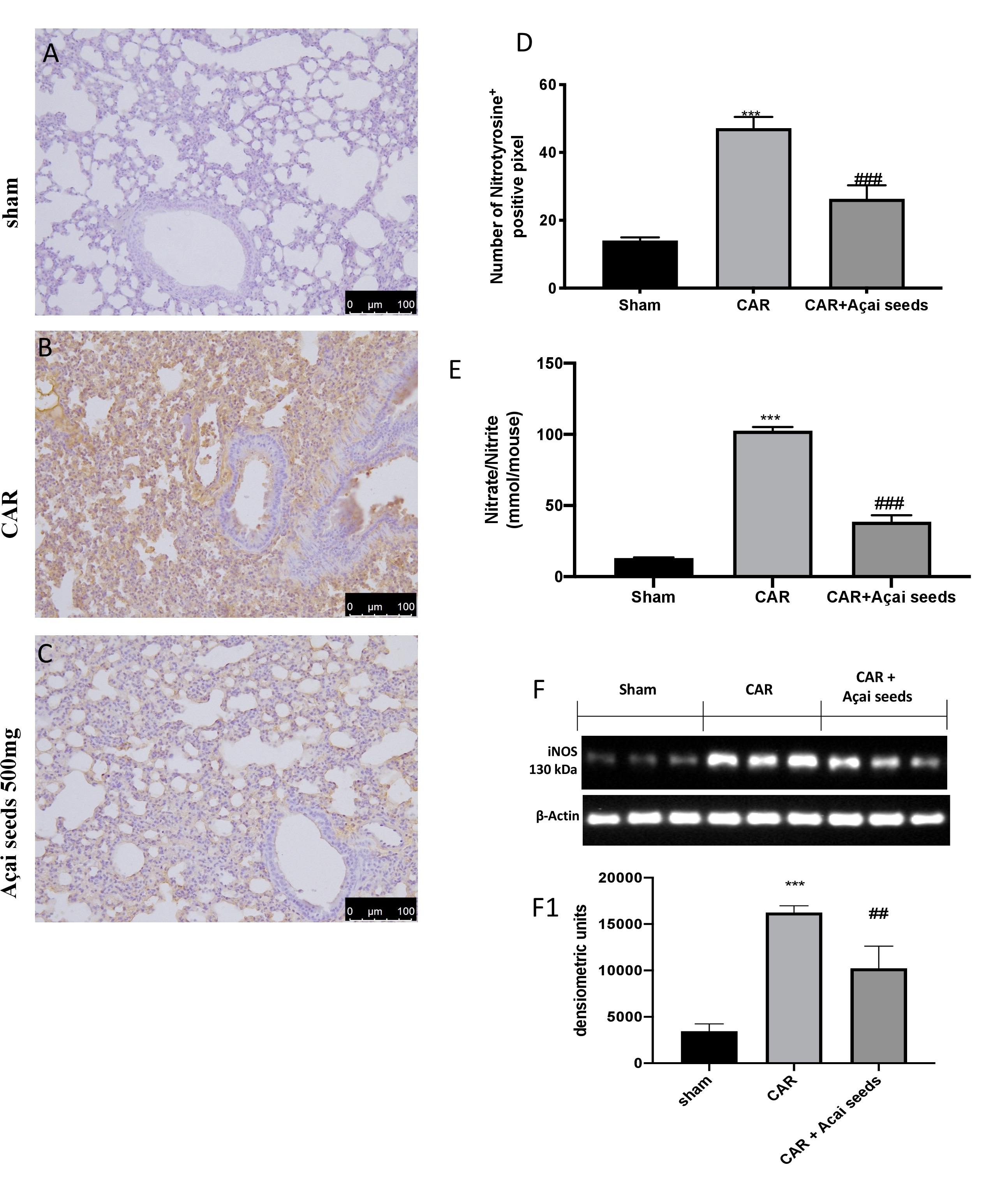
Açai Seeds reduce nitrosative stress CAR-induced
By western blots, immunohistochemistry and nitrite/nitrate analysis we investigated the effects of Açai Seeds administration on inhibition of nitrosative stress. We found that, after CAR injection there was a significantly increase in nitrotyrosine (Fig. 5B see graph 5D), iNOS (Fig. 5F see densitometric analysis F1) and nitrite/nitrate levels (Fig. 5E) compared to sham group (Fig. 5A see graph 5D for nitrotyrosine expression; Fig. 5F and densitometric analysis F1 for iNOS activity; Fig. 5E for nitrite/nitrate levels). As assumed, Açai Seeds administration protects the lung against nitrosative stress (Fig. 5A see graph 5D for nitrotyrosine expression; Fig. 5F and densitometric analisys F1 for iNOS activity; Fig. 5E for nitrite/nitrate levels).
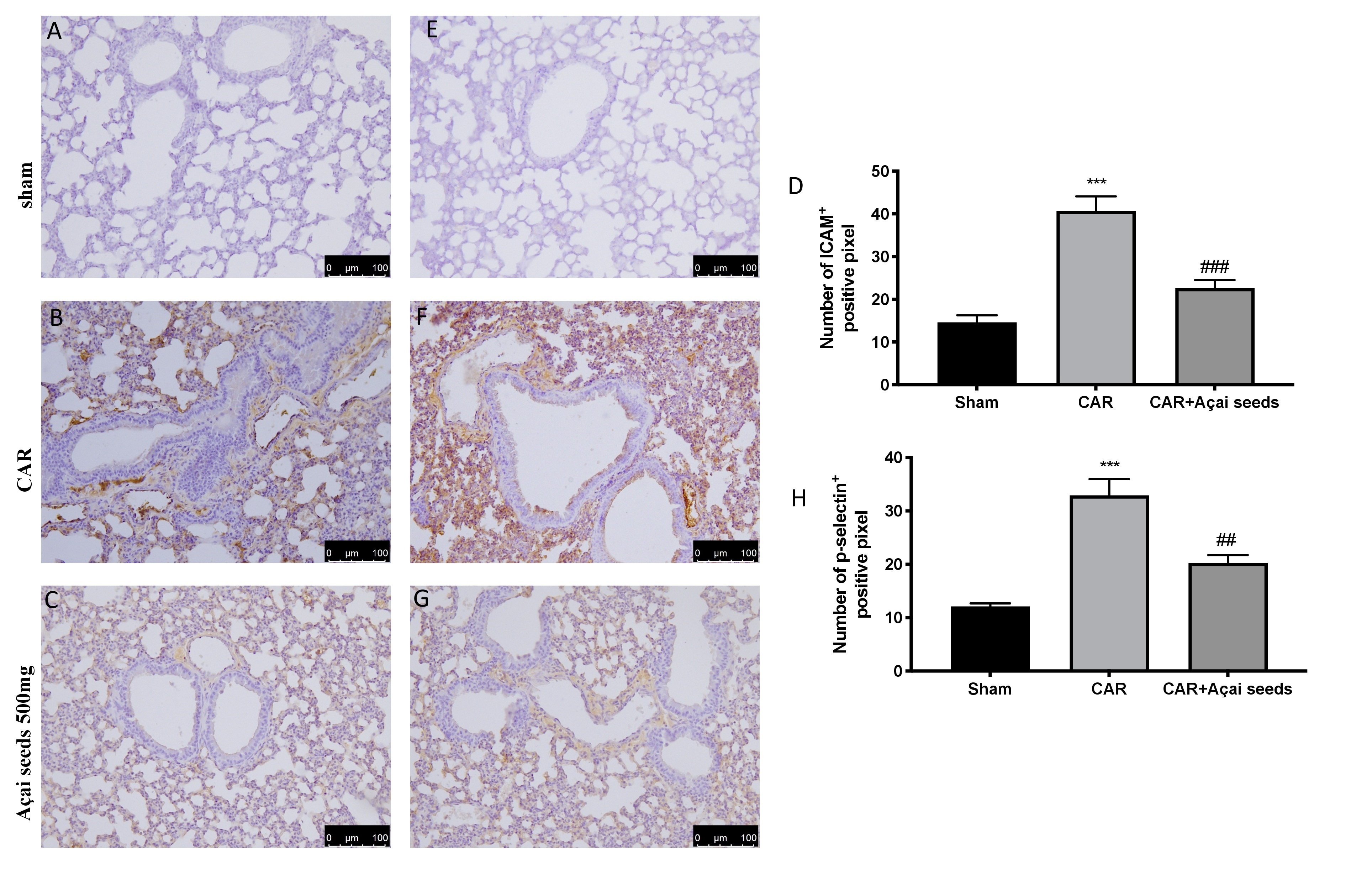
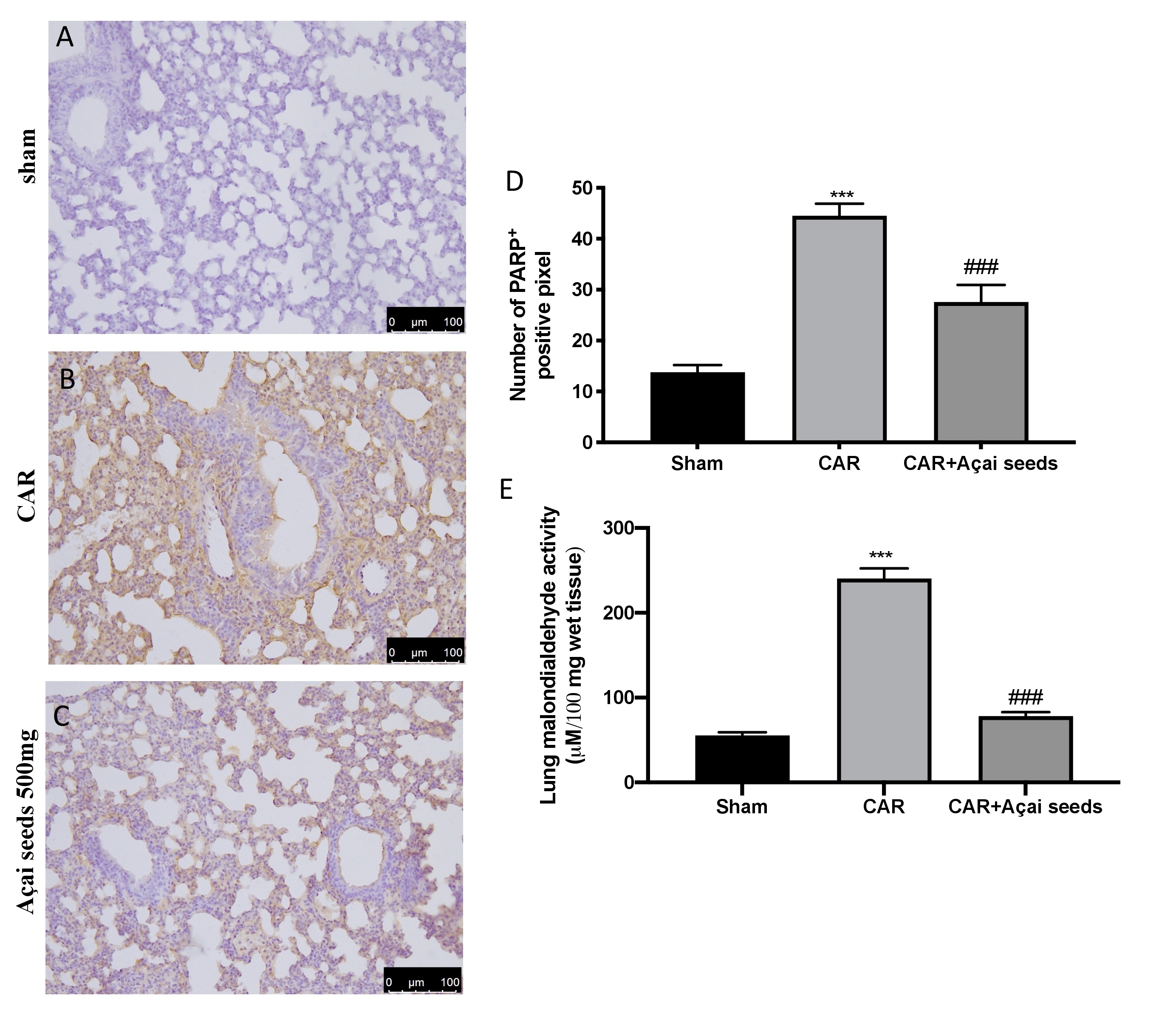
Açai Seeds administration reduce DNA damage and lipid peroxidation
Immunohistochemical analysis of lung tissue collected from CAR-treated group showed an increased in PARP positive cells (Fig. 7 Panel B, see graph D) compared to sham group (Fig. 7 Panel B, see graph D). Açai Seeds oral administration at the dose of 500 mg/kg was able to significantly decrease PARP expression (Fig. 6 Panel C, see graph D). Additionally, we evaluated lipid peroxidation by MDA activity, and we found that CAR injection in pleural cavity significantly increased lipid peroxidation compared to sham group (Fig. 7E). On the other hand, Açai Seeds oral administration at the dose of 500 mg/kg significantly decrease lipid peroxidation levels.
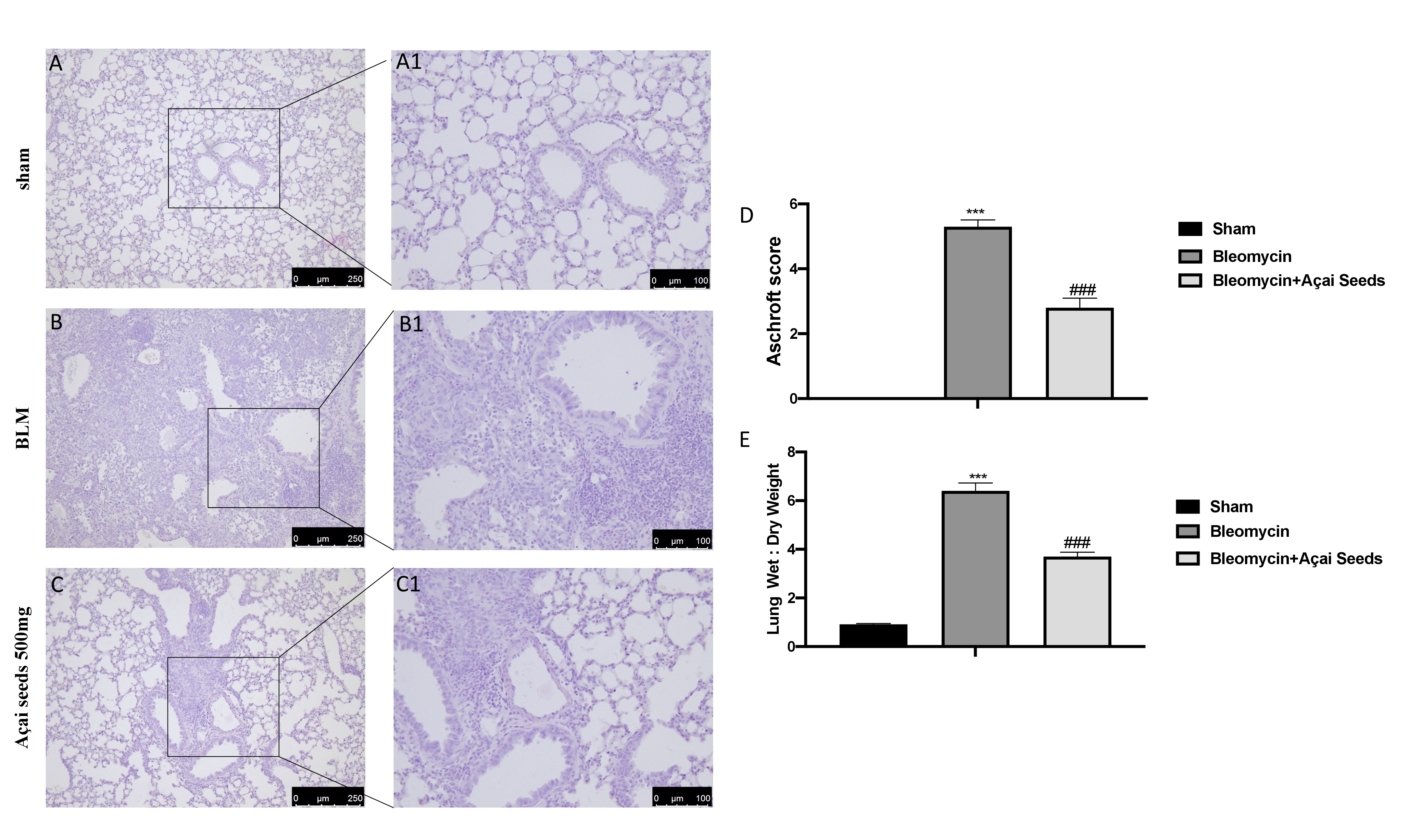
Açai Seeds reduce bleomycin-induced histological damage
Administration of bleomycin induce a significant alteration in the lung architecture, as highlighted in inflamed areas with notable presence of neutrophils and thickening of the lung tissue (Fig. 8B-B1 see Ashcroft score 8D) compared to the sham group (Fig. 8A-A1 see Ashcroft score 8D). Daily administration of Açai Seeds at the dose of 500 mg/kg for 14 days significantly decreases histological damage bleomycin-induced (Fig. 8C-C1 see Ashcroft score 8D). Additionally, the increase in the wet:dry lung weight ratio due to infiltration of inflammatory cells and edema founded in bleomycin group where significantly reduced after Açai Seeds administration (Fig. 1F).
Açai Seeds administration reduced bleomycin-induced inflammatory cell migration
Bleomycin damage is characterized by damage to the endothelial lining of small vessels and capillaries, accompanied by vascular congestion and increased microvascular permeability, which leads to an inflammatory response [79]. After bleomycin injection, we found an increase in cellular density in BAL (Fig. 9 from A to E for total cells, neutrophils, lymphocytes, and macrophages, respectively) as well as an increase in protein content (Fig. 9E) compared to the sham group. After 14 days of oral administration of Açai Seeds at the dose of 500 mg/kg we found a significantly decrease in all parameters evaluated.
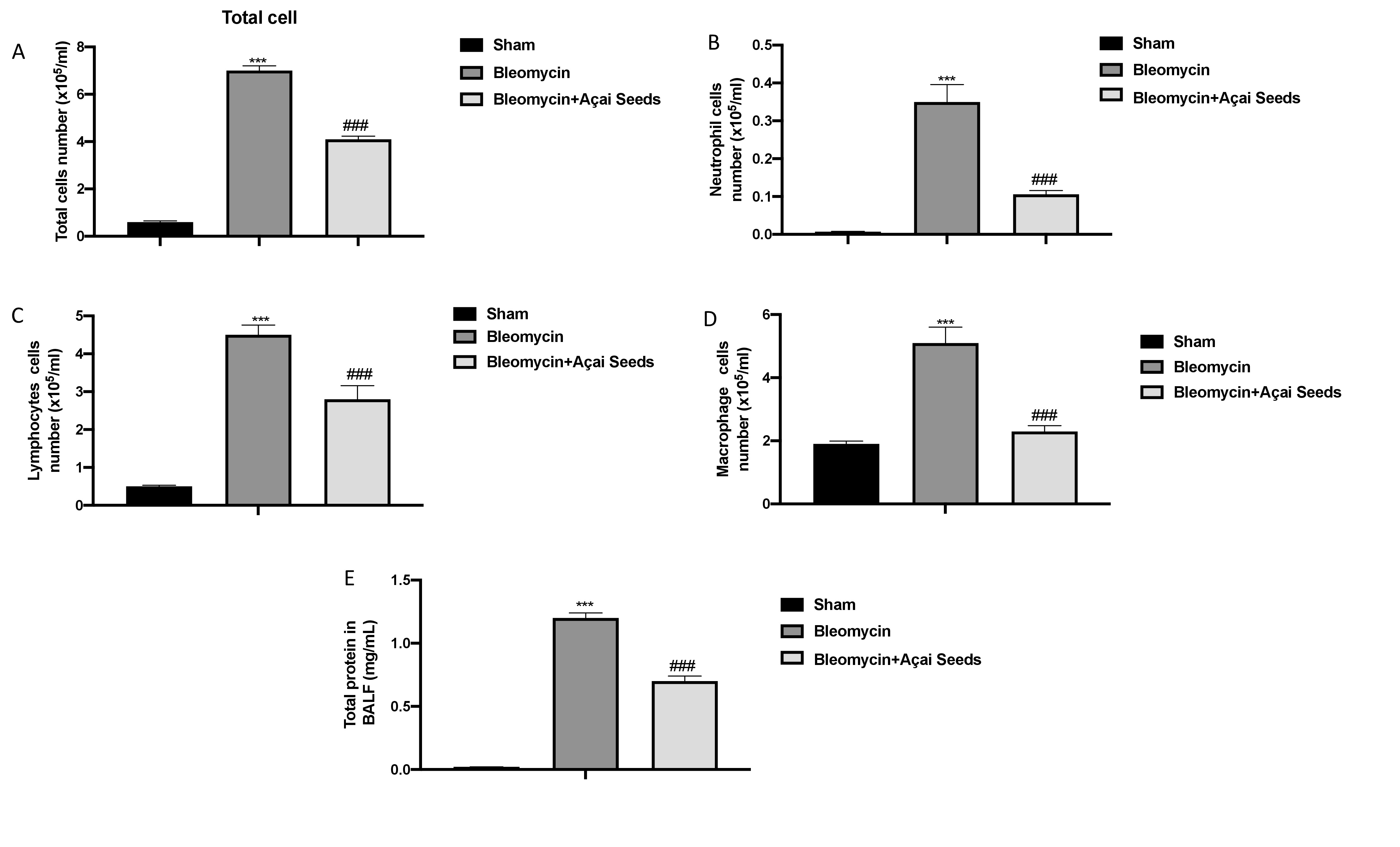
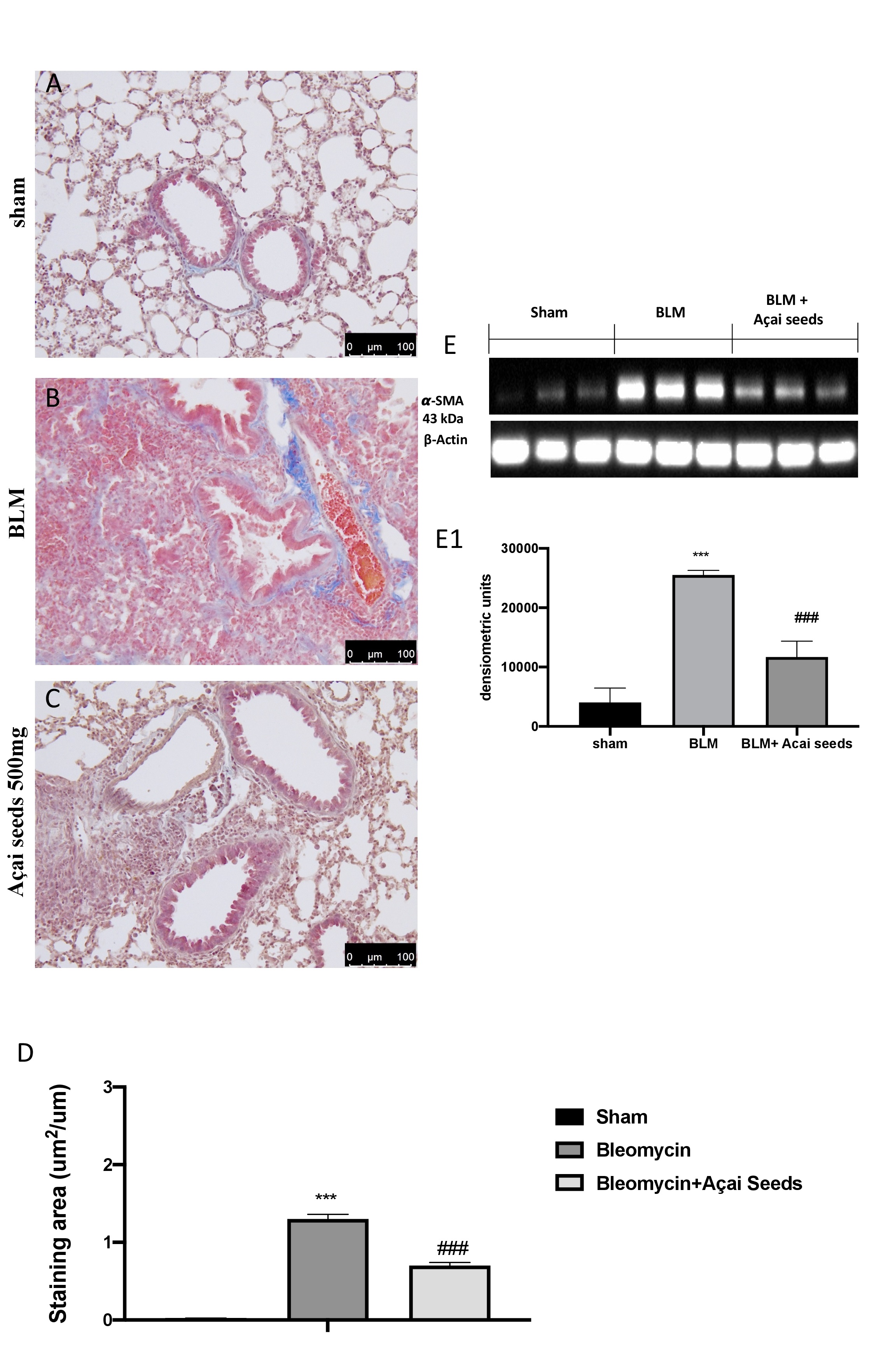
Açai Seeds decrease bleomycin-induced fibrosis
Masson’s trichrome staining and western blot analysis of a-SMA were used to evaluate the pulmonary fibrosis in lung tissue. After bleomycin injection, we found a significantly increase in fibrotic lesions and collagen accumulation in the lungs of mice (Fig. 10B and graph 10D) compared to the sham group (Fig. 10A and graph 10D). On the other hand, daily administration of Açai Seeds at the dose of 500 mg/kg significantly decreases collagen deposition and fibrotic scar. Another consolidated marker of fibrosis in the literature is represented by the expression of the a-SMA [30, 80]. As presumed, we found a significant increase in the group subjected to bleomycin when compared to the sham group (Fig. 10E and densitometric analysis 10-E1). A significantly decrease in a-SMA expression were found after Açai Seeds administration.
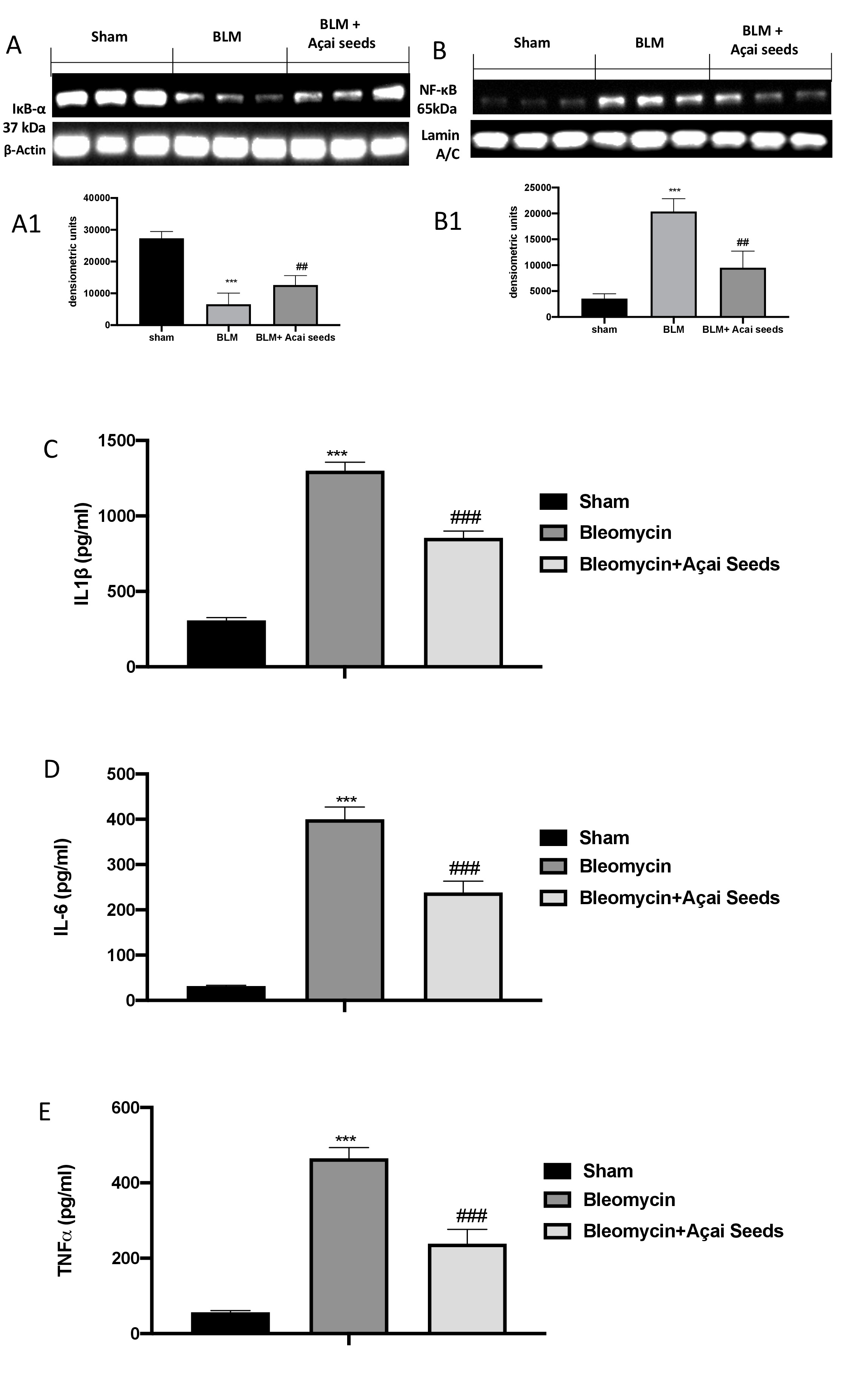
Açai Seeds administration decrease bleomycin-induced inflammation
To confirm our previous data on ALI, we investigated again by western blot the activation of NF-κB pathway. As assumed, we found a significantly decrease in IκB-a expression (Fig. 11A and A1), and consequently a significantly increase in NF-κB translocation (Fig. 11B and B1) in bleomycin group compared to sham group. On the other hand, we found a significantly restored in IκB-a expression (Fig. 11A and A1), and a significantly decrease in NF-κB translocation (Fig. 11B and B1). The same trend was observed in the analysis of IL-1b (Fig. 10C), IL-6 (Fig. 10D) and TNF-a (Fig. 10E).
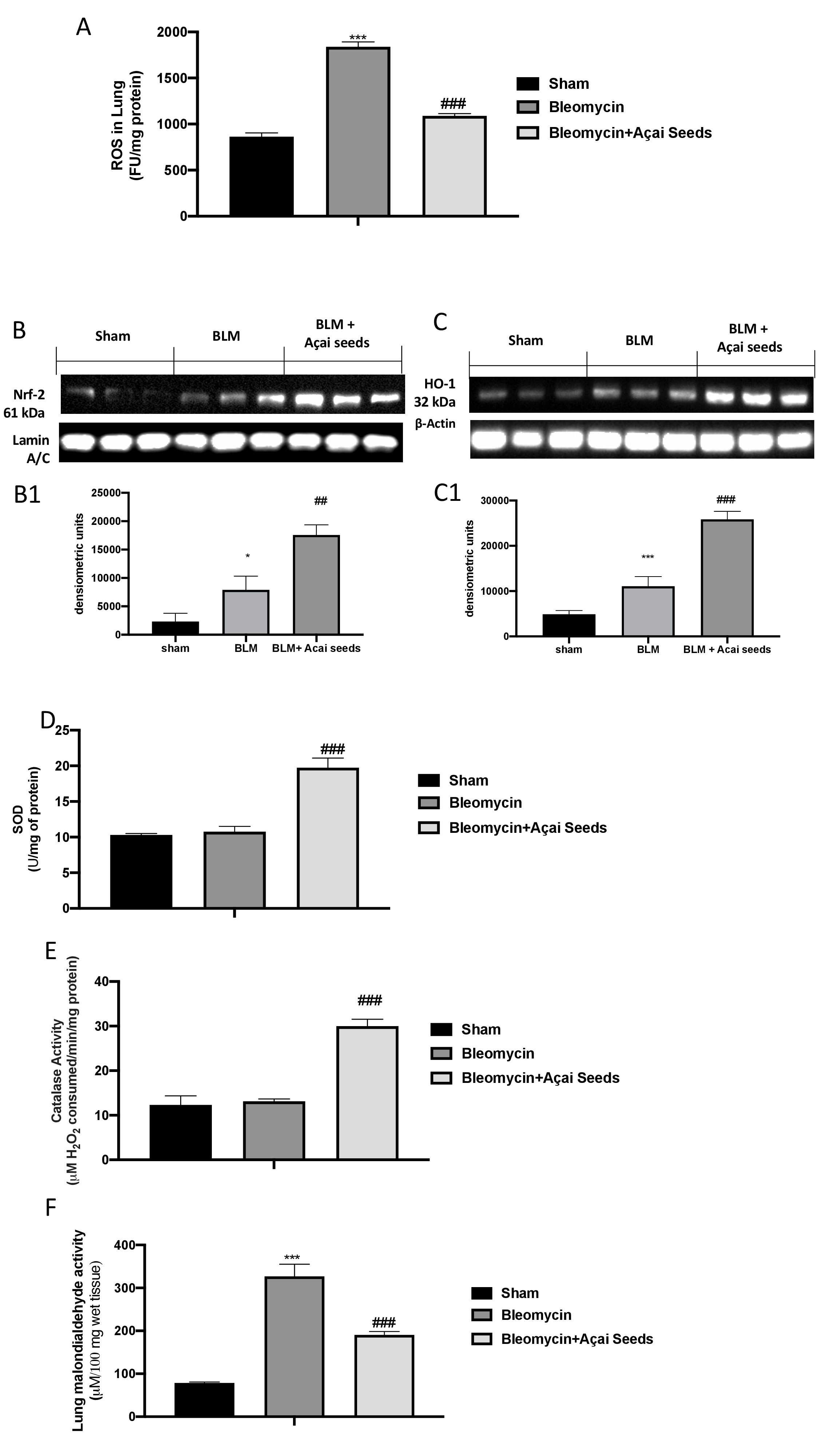
Açai Seeds administration increase physiological antioxidant defense and decrease lipid peroxidation in lung
As is well known, administration of bleomycin causes cellular toxicity via production of ROS, which leads to serious damage to the lungs (Fig.12A). By western blot we investigated Nrf-2 (Fig. 12B and B1) and HO-1 (Fig. 12C and C1) expressions, and we found that they were significantly increased after bleomycin injection compared to sham. After daily Açai Seeds administration at the dose of 500 mg/kg we found a significantly increase in Nrf-2 (Fig. 12B and B1) and HO-1 (Fig. 12C and C1) expressions. Considering that Nrf-2 is involved in the physiological regulation of the oxidative defense of the cells, we also investigated the activity of SOD (Fig. 11D) and CAT (Fig 11E). As assumed, we found that Açai Seeds administration was able to increase the physiological antioxidant response compared to bleomycin group. Therefore, in a similar way, the overproduction of free radicals resulted in an increase in lipid peroxidation, as reflected by the MDA levels induced by bleomycin administration, were significantly reduced after Açai Seeds administration (Fig. 11F).
Açai Seeds showed protection like Resveratrol
To compare the effects of Açai Seeds administration with the other well know antioxidants, we choose to make a new experiment with Resveratrol at the dose of 50 mg/kg. We found that the administration of Açai Seeds showed a significantly protection like resveratrol, reducing inflammation, architecture alteration, edema, and cells infiltration (Supplementary Fig. S1).
Discussion
In recent years, the use of natural compounds used to counter the onset of inflammatory events or even as a supplement to accompany drug treatments is enjoying great success [55, 56, 74, 81, 82]. Dietary supplements are in fact extensively utilized and have the potential to enhance health when given to people who need them. Although better diet quality is necessary to address these difficulties, dietary supplements and/or food fortification may be useful for those who are at risk of deficiency or have other medical disorders [83]. The Euterpe oleracea Mart. palm (Aceraceae), often known as “Açai” is a promising new source of natural antioxidants. This plant may be found across northern South America, having the highest concentration in Brazil’s Amazonian flood basins [19]. Despite the recent advances, Açai seeds warrant supplementary exploration as to their potential application for health benefits. Açai seeds extracts have previously demonstrated substantial antioxidant activity against the oxidation of linoleic acid as well as a potent scavenging ability against 2,2-diphenyl-1-picrylhydrazyl (DPPH) radicals and superoxide anion in a study of antioxidant activities of extracts from tropical and oriental medicinal plants [84]. Açai seeds extracts could thus provide benefits like grape seed or pine bark extracts, which are particularly high in oligomeric procyanidins. They have exhibited not only equivalent to or higher in vitro radical scavenging capability than butylated hydroxytoluene (BHT), but also cataract-prevention and antibacterial capabilities [19].
Lung damage, both acute and chronic, is a serious and rising worldwide health problem. The damage to many of these lung illnesses is irreversible, depleting the host’s ability to grow new lung, and existing medicines are just supportive rather than rehabilitative [85]. Both acute and chronic lung disease have in common several kinds of inflammatory cells that are activated during inflammation. Each one secretes cytokines and mediators that influence the behavior of other inflammatory cells. Inflammation progresses due to the orchestration of these cells and chemicals. Excessive inflammation in the lungs, which is a key organ for gas exchange, can be fatal. Because hazardous microorganisms are continually present in the lungs, a prompt and powerful defensive response is essential to eradicate the inflammation as quickly as possible [4].
In our experimental model of acute lung injury, we found a significantly alteration of lung architecture accompanied with an increase in cytokines release, mast cells degranulation, PMNs infiltration and MPO production after the injection in the pleural cavity of CAR. After the oral treatment with Açai Seeds we detected a considerably improvement in all these parameters.
Another key pathway involved during ALI development is oxidative and nitrosative stress. Phagocytic cells are the most prominent source of harmful oxidants. Although endothelial and alveolar epithelial cells may both produce oxidants, the predominance of oxidants in the lungs come from activated phagocytic cells [86, 87]. Nrf2, a critical nuclear transcriptional factor with significant antioxidative properties, has been frequently employed as a promoter to reduce oxidative stress and inflammation [88-91]. In our study, after CAR injection into the pleural cavity, we found a physiological increase in Nrf-2 expression as well as in HO-1, that were increased after Açai Seeds administration. As a consequence of oxidative stress, we found also a significantly increase in DNA damage and lipid peroxidation. Açai Seeds administration significantly reduce lipid peroxidation as well as DNA damage. Oxidative stress and the impairment in the physiological antioxidant response are a key pathway involved in bleomycin-induce lung injury. In our experiment after the induction of lung injury with an instillation of bleomycin we found a significantly increase in lung alteration due to an increase in fibrosis, as demonstrated with the increase in a-SMA expression. After 14 days of oral daily treatment with Açai Seeds, we notice a significantly decrease in histological damage as well as in fibrosis. As demonstrated in acute model, we also found that Açai Seeds administration was able to reduce inflammatory pathways and increase physiological endogenous response.
Conclusion
Our study demonstrated not only that Açai Seeds nutritional supplementation have a beneficial effect on acute and chronic lung disease but also that it’s probably act by the modulation of NF-kB and Nrf-2 pathways.
Author Contributions
Conceptualization, Marika Cordaro; Data curation, Alessio Peritore; Investigation, Livia Interdonato and Tiziana Genovese; Methodology, Ramona D’Amico, Rosalia Crupi and Enrico Gugliandolo; Project administration, Salvatore Cuzzocrea and Rosanna Di Paola; Supervision, Tiziana Genovese; Validation, Roberta Fusco and Daniela Impellizzeri; Writing – original draft, Marika Cordaro; Writing – review & editing, Rosalba Siracusa. All authors have read and agreed to the published version of the manuscript.
Statement of Ethics
The University of Messina Review Board for animal care (OPBA) approved the study (Ethical Code: 89126.24/2021-PR). All animal experiments agree with the new Italian regulations (D.Lgs 2014/26), EU regulations (EU Directive 2010/63) and the ARRIVE guidelines.
The authors declare that no conflicts of interest exist.
| 1 Ware LB, Matthay MA: The acute respiratory distress syndrome. N Engl J Med 2000;342:1334-1349. https://doi.org/10.1056/NEJM200005043421806 |
||||
| 2 Matthay MA, Zimmerman GA: Acute lung injury and the acute respiratory distress syndrome: four decades of inquiry into pathogenesis and rational management. Am J Respir Cell Mol Biol 2005;33:319-327. https://doi.org/10.1165/rcmb.F305 |
||||
| 3 Maybauer MO, Maybauer DM, Herndon DN: Incidence and outcomes of acute lung injury. N Engl J Med 2006;354:416-417; author reply 416-417. https://doi.org/10.1056/NEJMc053159 |
||||
| 4 Moldoveanu B, Otmishi P, Jani P, Walker J, Sarmiento X, Guardiola J, Saad M, Yu J: Inflammatory mechanisms in the lung. J Inflamm Res 2009;2:1-11. https://doi.org/10.4137/CCRPM.S547 |
||||
| 5 Barnes PJ, Shapiro SD, Pauwels RA: Chronic obstructive pulmonary disease: molecular and cellular mechanisms. Eur Respir J 2003;22:672-688. https://doi.org/10.1183/09031936.03.00040703 |
||||
| 6 Rahman I: Oxidative stress in pathogenesis of chronic obstructive pulmonary disease: cellular and molecular mechanisms. Cell Biochem Biophys 2005;43:167-188. https://doi.org/10.1385/CBB:43:1:167 |
||||
| 7 Sykiotis GP, Bohmann D: Stress-activated cap'n'collar transcription factors in aging and human disease. Sci Signal 2010;3:re3. https://doi.org/10.1126/scisignal.3112re3 |
||||
| 8 Kensler TW, Wakabayashi N, Biswal S: Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol 2007;47:89-116. https://doi.org/10.1146/annurev.pharmtox.46.120604.141046 |
||||
| 9 Zhang DD: Mechanistic studies of the Nrf2-Keap1 signaling pathway. Drug Metab Rev 2006;38:769-789. https://doi.org/10.1080/03602530600971974 |
||||
| 10 Carlson J, Price L, Deng H: Nrf2 and the Nrf2-interacting network in respiratory inflammation and diseases. Nrf2 and its Modulation in Inflammation 2020;85:51-76. https://doi.org/10.1007/978-3-030-44599-7_3 |
||||
| 11 Malhotra D, Portales-Casamar E, Singh A, Srivastava S, Arenillas D, Happel C, Shyr C, Wakabayashi N, Kensler TW, Wasserman WW, Biswal S: Global mapping of binding sites for Nrf2 identifies novel targets in cell survival response through ChIP-Seq profiling and network analysis. Nucleic Acids Res 2010;38:5718-5734. https://doi.org/10.1093/nar/gkq212 |
||||
| 12 Lingappan K: NF-kappaB in Oxidative Stress. Curr Opin Toxicol 2018;7:81-86. https://doi.org/10.1016/j.cotox.2017.11.002 |
||||
| 13 de Boer A, van de Worp W, Hageman GJ, Bast A: The effect of dietary components on inflammatory lung diseases - a literature review. Int J Food Sci Nutr 2017;68:771-787. https://doi.org/10.1080/09637486.2017.1288199 |
||||
| 14 Johnson ER, Matthay MA: Acute lung injury: epidemiology, pathogenesis, and treatment. J Aerosol Med Pulm Drug Deliv 2010;23:243-252. https://doi.org/10.1089/jamp.2009.0775 |
||||
| 15 Wood LG, Shivappa N, Berthon BS, Gibson PG, Hebert JR: Dietary inflammatory index is related to asthma risk, lung function and systemic inflammation in asthma. Clin Exp Allergy 2015;45:177-183. https://doi.org/10.1111/cea.12323 |
||||
| 16 Maciel‐Silva FW, Buller LS, Gonçalves MLMBB, Rostagno MA, Forster‐Carneiro T: Sustainable development in the Legal Amazon: energy recovery from açaí seeds. Biofuels Bioprod Bioref 2021;15:1174-1189. https://doi.org/10.1002/bbb.2222 |
||||
| 17 Melo PS, Massarioli AP, Lazarini JG, Soares JC, Franchin M, Rosalen PL, Alencar SM: Simulated gastrointestinal digestion of Brazilian acai seeds affects the content of flavan-3-ol derivatives, and their antioxidant and anti-inflammatory activities. Heliyon 2020;6:e05214. https://doi.org/10.1016/j.heliyon.2020.e05214 |
||||
| 18 Melo PS, Selani MM, Gonçalves RH, de Oliveira Paulino J, Massarioli AP, de Alencar SM: Açaí seeds: An unexplored agro-industrial residue as a potential source of lipids, fibers, and antioxidant phenolic compounds. Ind Crops Prod 2021;161:113204. https://doi.org/10.1016/j.indcrop.2020.113204 |
||||
| 19 Rodrigues RB, Lichtenthaler R, Zimmermann BF, Papagiannopoulos M, Fabricius H, Marx F, Maia JG, Almeida O: Total oxidant scavenging capacity of Euterpe oleracea Mart. (acai) seeds and identification of their polyphenolic compounds. J Agric Food Chem 2006;54:4162-4167. https://doi.org/10.1021/jf058169p |
||||
| 20 de Moura RS, Pires KM, Santos Ferreira T, Lopes AA, Nesi RT, Resende AC, Sousa PJ, da Silva AJ, Porto LC, Valenca SS: Addition of acai (Euterpe oleracea) to cigarettes has a protective effect against emphysema in mice. Food Chem Toxicol 2011;49:855-863. https://doi.org/10.1016/j.fct.2010.12.007 |
||||
| 21 Lee JY, Kim N, Choi YJ, Nam RH, Lee S, Ham MH, Suh JH, Choi YJ, Lee HS, Lee DH: Anti-inflammatory and Anti-tumorigenic Effects of Acai Berry in Helicobacter felis-infected mice. J Cancer Prev 2016;21:48-54. https://doi.org/10.15430/JCP.2016.21.1.48 |
||||
| 22 Moura RS, Ferreira TS, Lopes AA, Pires KM, Nesi RT, Resende AC, Souza PJ, Silva AJ, Borges RM, Porto LC, Valenca SS: Effects of Euterpe oleracea Mart. (ACAI) extract in acute lung inflammation induced by cigarette smoke in the mouse. Phytomedicine 2012;19:262-269. https://doi.org/10.1016/j.phymed.2011.11.004 |
||||
| 23 Poulose SM, Fisher DR, Larson J, Bielinski DF, Rimando AM, Carey AN, Schauss AG, Shukitt-Hale B: Anthocyanin-rich acai (Euterpe oleracea Mart.) fruit pulp fractions attenuate inflammatory stress signaling in mouse brain BV-2 microglial cells. J Agric Food Chem 2012;60:1084-1093. https://doi.org/10.1021/jf203989k |
||||
| 24 Santos IB, de Bem GF, da Costa CA, de Carvalho L, de Medeiros AF, Silva DLB, Romao MH, de Andrade Soares R, Ognibene DT, de Moura RS, Resende AC: Acai seed extract prevents the renin-angiotensin system activation, oxidative stress and inflammation in white adipose tissue of high-fat diet-fed mice. Nutr Res 2020;79:35-49. https://doi.org/10.1016/j.nutres.2020.05.006 |
||||
| 25 Silva DF, Vidal FC, Santos D, Costa MC, Morgado-Diaz JA, do Desterro Soares Brandao Nascimento M, de Moura RS: Cytotoxic effects of Euterpe oleracea Mart. in malignant cell lines. BMC Complement Altern Med 2014;14:175. https://doi.org/10.1186/1472-6882-14-175 |
||||
| 26 Barros L, Calhelha RC, Queiroz MJR, Santos-Buelga C, Santos EA, Regis WC, Ferreira IC: The powerful in vitro bioactivity of Euterpe oleracea Mart. seeds and related phenolic compounds. Ind Crops Prod 2015;76:318-322. https://doi.org/10.1016/j.indcrop.2015.05.086 |
||||
| 27 Di Paola R, Mazzon E, Muia C, Genovese T, Menegazzi M, Zaffini R, Suzuki H, Cuzzocrea S: Green tea polyphenol extract attenuates lung injury in experimental model of carrageenan-induced pleurisy in mice. Respir Res 2005;6:66. https://doi.org/10.1186/1465-9921-6-66 |
||||
| 28 da Silva RC, Batista A, Costa D, Moura-Nunes N, Koury JC, da Costa CA, Resende AC, Daleprane JB: Acai (Euterpe oleracea Mart.) seed flour prevents obesity-induced hepatic steatosis regulating lipid metabolism by increasing cholesterol excretion in high-fat diet-fed mice. Food Res Int 2018;111:408-415. https://doi.org/10.1016/j.foodres.2018.05.043 |
||||
| 29 Impellizzeri D, Talero E, Siracusa R, Alcaide A, Cordaro M, Maria Zubelia J, Bruschetta G, Crupi R, Esposito E, Cuzzocrea S, Motilva V: Protective effect of polyphenols in an inflammatory process associated with experimental pulmonary fibrosis in mice. Br J Nutr 2015;114:853-865. https://doi.org/10.1017/S0007114515002597 |
||||
| 30 Conte E, Fagone E, Gili E, Fruciano M, Iemmolo M, Pistorio MP, Impellizzeri D, Cordaro M, Cuzzocrea S, Vancheri C: Preventive and therapeutic effects of thymosin beta4 N-terminal fragment Ac-SDKP in the bleomycin model of pulmonary fibrosis. Oncotarget 2016;7:33841-33854. https://doi.org/10.18632/oncotarget.8409 |
||||
| 31 Fusco R, Cordaro M, Genovese T, Impellizzeri D, Siracusa R, Gugliandolo E, Peritore AF, D'Amico R, Crupi R, Cuzzocrea S, Di Paola R: Adelmidrol: A New Promising Antioxidant and Anti-Inflammatory Therapeutic Tool in Pulmonary Fibrosis. Antioxidants (Basel) 2020;9:601. https://doi.org/10.3390/antiox9070601 |
||||
| 32 Mariotto S, Esposito E, Di Paola R, Ciampa A, Mazzon E, Carcereri de Prati A, Darra E, Vincenzi S, Cucinotta G, Caminiti R, Suzuki H, Cuzzocrea S: Protective effect of Arbutus unedo aqueous extract in carrageenan-induced lung inflammation in mice. Pharmacol Res 2008;57:110-124. https://doi.org/10.1016/j.phrs.2007.12.005 |
||||
| 33 Vaughan RP, Szewczyk MT, Jr., Lanosa MJ, Desesa CR, Gianutsos G, Morris JB: Adenosine sensory transduction pathways contribute to activation of the sensory irritation response to inspired irritant vapors. Toxicol Sci 2006;93:411-421. https://doi.org/10.1093/toxsci/kfl061 |
||||
| 34 D'Amico R, Monaco F, Fusco R, Peritore AF, Genovese T, Impellizzeri D, Crupi R, Interdonato L, Sforza AM, Gugliandolo E, Siracusa R, Cuzzocrea S, Cordaro M, Di Paola R: Exposure to Atrazine Induces Lung Inflammation through Nrf2-HO1 and Beclin 1/LC3 Pathways. Cell Physiol Biochem 2021;55:413-427. https://doi.org/10.33594/000000393 |
||||
| 35 Lin J, Xia J, Zhao HS, Hou R, Talukder M, Yu L, Guo JY, Li JL: Lycopene Triggers Nrf2-AMPK Cross Talk to Alleviate Atrazine-Induced Nephrotoxicity in Mice. J Agric Food Chem 2018;66:12385-12394. https://doi.org/10.1021/acs.jafc.8b04341 |
||||
| 36 Liu W, Du Y, Liu J, Wang H, Sun D, Liang D, Zhao L, Shang J: Effects of atrazine on the oxidative damage of kidney in Wister rats. Int J Clin Exp Med 2014;7:3235-3243. | ||||
| 37 Zhang X, Wang M, Gao S, Ren R, Zheng J, Zhang Y: Atrazine-induced apoptosis of splenocytes in BALB/C mice. BMC Med 2011;9:117. https://doi.org/10.1186/1741-7015-9-117 |
||||
| 38 Li J, Li X, Bi H, Ma K, Li B: Developmental Exposure to Atrazine Impairs Spatial Memory and Downregulates the Hippocampal D1 Dopamine Receptor and cAMP-Dependent Signaling Pathway in Rats. Int J Mol Sci 2018;19:2241. https://doi.org/10.3390/ijms19082241 |
||||
| 39 Gao S, Wang Z, Zhang C, Jia L, Zhang Y: Oral Exposure to Atrazine Induces Oxidative Stress and Calcium Homeostasis Disruption in Spleen of Mice. Oxid Med Cell Longev 2016;2016:7978219. https://doi.org/10.1155/2016/7978219 |
||||
| 40 Schramm CM, Puddington L, Wu C, Guernsey L, Gharaee-Kermani M, Phan SH, Thrall RS: Chronic inhaled ovalbumin exposure induces antigen-dependent but not antigen-specific inhalational tolerance in a murine model of allergic airway disease. Am J Pathol 2004;164:295-304. https://doi.org/10.1016/S0002-9440(10)63119-7 |
||||
| 41 Fusco R, Gugliandolo E, Biundo F, Campolo M, Di Paola R, Cuzzocrea S: Inhibition of inflammasome activation improves lung acute injury induced by carrageenan in a mouse model of pleurisy. FASEB J 2017;31:3497-3511. https://doi.org/10.1096/fj.201601349R |
||||
| 42 Di Paola R, Impellizzeri D, Fusco R, Cordaro M, Siracusa R, Crupi R, Esposito E, Cuzzocrea S: Ultramicronized palmitoylethanolamide (PEA-um((R))) in the treatment of idiopathic pulmonary fibrosis. Pharmacol Res 2016;111:405-412. https://doi.org/10.1016/j.phrs.2016.07.010 |
||||
| 43 Manitsopoulos N, Nikitopoulou I, Maniatis NA, Magkou C, Kotanidou A, Orfanos SE: Highly Selective Endothelin-1 Receptor A Inhibition Prevents Bleomycin-Induced Pulmonary Inflammation and Fibrosis in Mice. Respiration 2018;95:122-136. https://doi.org/10.1159/000481201 |
||||
| 44 Kato S, Inui N, Hakamata A, Suzuki Y, Enomoto N, Fujisawa T, Nakamura Y, Watanabe H, Suda T: Changes in pulmonary endothelial cell properties during bleomycin-induced pulmonary fibrosis. Respir Res 2018;19:127. https://doi.org/10.1186/s12931-018-0831-y |
||||
| 45 Cordaro M, Paterniti I, Siracusa R, Impellizzeri D, Esposito E, Cuzzocrea S: KU0063794, a Dual mTORC1 and mTORC2 Inhibitor, Reduces Neural Tissue Damage and Locomotor Impairment After Spinal Cord Injury in Mice. Mol Neurobiol 2017;54:2415-2427. https://doi.org/10.1007/s12035-016-9827-0 |
||||
| 46 Siracusa R, Impellizzeri D, Cordaro M, Crupi R, Esposito E, Petrosino S, Cuzzocrea S: Anti-Inflammatory and Neuroprotective Effects of Co-UltraPEALut in a Mouse Model of Vascular Dementia. Front Neurol 2017;8:233. https://doi.org/10.3389/fneur.2017.00233 |
||||
| 47 Campolo M, Esposito E, Ahmad A, Di Paola R, Paterniti I, Cordaro M, Bruschetta G, Wallace JL, Cuzzocrea S: Hydrogen sulfide-releasing cyclooxygenase inhibitor ATB-346 enhances motor function and reduces cortical lesion volume following traumatic brain injury in mice. J Neuroinflammation 2014;11:196. https://doi.org/10.1186/s12974-014-0196-1 |
||||
| 48 Paterniti I, Di Paola R, Campolo M, Siracusa R, Cordaro M, Bruschetta G, Tremolada G, Maestroni A, Bandello F, Esposito E, Zerbini G, Cuzzocrea S: Palmitoylethanolamide treatment reduces retinal inflammation in streptozotocin-induced diabetic rats. Eur J Pharmacol 2015;769:313-323. https://doi.org/10.1016/j.ejphar.2015.11.035 |
||||
| 49 Cordaro M, Impellizzeri D, Gugliandolo E, Siracusa R, Crupi R, Esposito E, Cuzzocrea S: Adelmidrol, a Palmitoylethanolamide Analogue, as a New Pharmacological Treatment for the Management of Inflammatory Bowel Disease. Mol Pharmacol 2016;90:549-561. https://doi.org/10.1124/mol.116.105668 |
||||
| 50 Di Paola R, Cordaro M, Crupi R, Siracusa R, Campolo M, Bruschetta G, Fusco R, Pugliatti P, Esposito E, Cuzzocrea S: Protective Effects of Ultramicronized Palmitoylethanolamide (PEA-um) in Myocardial Ischaemia and Reperfusion Injury in vivo . Shock 2016;46:202-213. https://doi.org/10.1097/SHK.0000000000000578 |
||||
| 51 Esposito E, Impellizzeri D, Bruschetta G, Cordaro M, Siracusa R, Gugliandolo E, Crupi R, Cuzzocrea S: A new co-micronized composite containing palmitoylethanolamide and polydatin shows superior oral efficacy compared to their association in a rat paw model of carrageenan-induced inflammation. Eur J Pharmacol 2016;782:107-118. https://doi.org/10.1016/j.ejphar.2016.03.033 |
||||
| 52 Impellizzeri D, Cordaro M, Bruschetta G, Crupi R, Pascali J, Alfonsi D, Marcolongo G, Cuzzocrea S: 2-pentadecyl-2-oxazoline: Identification in coffee, synthesis and activity in a rat model of carrageenan-induced hindpaw inflammation. Pharmacol Res 2016;108:23-30. https://doi.org/10.1016/j.phrs.2016.04.007 |
||||
| 53 Fusco R, D'Amico R, Cordaro M, Gugliandolo E, Siracusa R, Peritore AF, Crupi R, Impellizzeri D, Cuzzocrea S, Di Paola R: Absence of formyl peptide receptor 1 causes endometriotic lesion regression in a mouse model of surgically-induced endometriosis. Oncotarget 2018;9:31355-31366. https://doi.org/10.18632/oncotarget.25823 |
||||
| 54 Gugliandolo E, D'Amico R, Cordaro M, Fusco R, Siracusa R, Crupi R, Impellizzeri D, Cuzzocrea S, Di Paola R: Effect of PEA-OXA on neuropathic pain and functional recovery after sciatic nerve crush. J Neuroinflammation 2018;15:264. https://doi.org/10.1186/s12974-018-1303-5 |
||||
| 55 Fusco R, Siracusa R, Peritore AF, Gugliandolo E, Genovese T, D'Amico R, Cordaro M, Crupi R, Mandalari G, Impellizzeri D, Cuzzocrea S, Di Paola R: The Role of Cashew (Anacardium occidentale L.) Nuts on an Experimental Model of Painful Degenerative Joint Disease. Antioxidants (Basel) 2020;9:511. https://doi.org/10.3390/antiox9060511 |
||||
| 56 Siracusa R, Fusco R, Peritore AF, Cordaro M, D'Amico R, Genovese T, Gugliandolo E, Crupi R, Smeriglio A, Mandalari G, Cuzzocrea S, Di Paola R, Impellizzeri D: The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020;12:834. https://doi.org/10.3390/nu12030834 |
||||
| 57 Di Paola R, Fusco R, Impellizzeri D, Cordaro M, Britti D, Morittu VM, Evangelista M, Cuzzocrea S: Adelmidrol, in combination with hyaluronic acid, displays increased anti-inflammatory and analgesic effects against monosodium iodoacetate-induced osteoarthritis in rats. Arthritis Res Ther 2016;18:291. https://doi.org/10.1186/s13075-016-1189-5 |
||||
| 58 Di Paola R, Crisafulli C, Mazzon E, Genovese T, Paterniti I, Bramanti P, Cuzzocrea S: Effect of PD98059, a selective MAPK3/MAPK1 inhibitor, on acute lung injury in mice. Int J Immunopathol Pharmacol 2009;22:937-950. https://doi.org/10.1177/039463200902200409 |
||||
| 59 Liang J, Wu S, Xie W, He H: Ketamine ameliorates oxidative stress-induced apoptosis in experimental traumatic brain injury via the Nrf2 pathway. Drug Des Devel Ther 2018;12:845-853. https://doi.org/10.2147/DDDT.S160046 |
||||
| 60 Zhang R, Liu C, Li Y, Chen L, Xiang J: Tenacissoside H Promotes Neurological Recovery of Cerebral Ischemia-reperfusion Injury in Mice by Modulating Inflammation and Oxidative stress via TrkB Pathway. Clin Exp Pharmacol Physiol 2021;48:757-769. https://doi.org/10.1111/1440-1681.13398 |
||||
| 61 Shi DD, Huang YH, Lai CSW, Dong CM, Ho LC, Wu EX, Li Q, Wang XM, Chung SK, Sham PC, Zhang ZJ: Chemotherapy-Induced Cognitive Impairment Is Associated with Cytokine Dysregulation and Disruptions in Neuroplasticity. Mol Neurobiol 2019;56:2234-2243. https://doi.org/10.1007/s12035-018-1224-4 |
||||
| 62 Zhu N, Liang X, Zhang M, Yin X, Yang H, Zhi Y, Ying G, Zou J, Chen L, Yao X, Li H: Astaxanthin protects cognitive function of vascular dementia. Behav Brain Funct 2020;16:10. https://doi.org/10.1186/s12993-020-00172-8 |
||||
| 63 Peritore AF, Siracusa R, Fusco R, Gugliandolo E, D'Amico R, Cordaro M, Crupi R, Genovese T, Impellizzeri D, Cuzzocrea S, Di Paola R: Ultramicronized Palmitoylethanolamide and Paracetamol, a New Association to Relieve Hyperalgesia and Pain in a Sciatic Nerve Injury Model in Rat. Int J Mol Sci 2020;21:3509. https://doi.org/10.3390/ijms21103509 |
||||
| 64 An X, Sun X, Hou Y, Yang X, Chen H, Zhang P, Wu J: Protective effect of oxytocin on LPS-induced acute lung injury in mice. Sci Rep 2019;9:2836. https://doi.org/10.1038/s41598-019-39349-1 |
||||
| 65 Marklund S, Marklund G: Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem 1974;47:469-474. https://doi.org/10.1111/j.1432-1033.1974.tb03714.x |
||||
| 66 Aebi H: Catalase in vitro . Methods Enzymol 1984;105:121-126. https://doi.org/10.1016/S0076-6879(84)05016-3 |
||||
| 67 Moron MS, Depierre JW, Mannervik B: Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim Biophys Acta 1979;582:67-78. https://doi.org/10.1016/0304-4165(79)90289-7 |
||||
| 68 Arora A, Bhuria V, Hazari PP, Pathak U, Mathur S, Roy BG, Sandhir R, Soni R, Dwarakanath BS, Bhatt AN: Amifostine Analog, DRDE-30, Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Front Pharmacol 2018;9:394. https://doi.org/10.3389/fphar.2018.00394 |
||||
| 69 Fusco R, Salinaro AT, Siracusa R, D'Amico R, Impellizzeri D, Scuto M, Ontario ML, Crea R, Cordaro M, Cuzzocrea S: Hidrox® Counteracts Cyclophosphamide-Induced Male Infertility through NRF2 Pathways in a Mouse Model. Antioxidants (Basel) 2021;10:778. https://doi.org/10.3390/antiox10050778 |
||||
| 70 Lebel CP, Ali SF, Mckee M, Bondy SC: Organometal-Induced Increases in Oxygen Reactive Species - the Potential of 2',7'-Dichlorofluorescin Diacetate as an Index of Neurotoxic Damage. Toxicol Appl Pharmacol 1990;104:17-24. https://doi.org/10.1016/0041-008X(90)90278-3 |
||||
| 71 Nemmar A, Raza H, Yuvaraju P, Beegam S, John A, Yasin J, Hameed RS, Adeghate E, Ali BH: Nose-only water-pipe smoking effects on airway resistance, inflammation, and oxidative stress in mice. J Appl Physiol 2013;115:1316-1323. https://doi.org/10.1152/japplphysiol.00194.2013 |
||||
| 72 Nemmar A, Karaca T, Beegam S, Yuvaraju P, Yasin J, Ali BH: Lung Oxidative Stress, DNA Damage, Apoptosis, and Fibrosis in Adenine-Induced Chronic Kidney Disease in Mice. Front Physiol 2017;8:896. https://doi.org/10.3389/fphys.2017.00896 |
||||
| 73 D'Amico R, Monaco F, Fusco R, Peritore AF, Genovese T, Impellizzeri D, Crupi R, Interdonato L, Sforza AM, Gugliandolo E: Exposure to Atrazine Induces Lung Inflammation through Nrf2-HO1 and Beclin 1/LC3 Pathways. Cell Physiol Biochem 2021;55:413-427. https://doi.org/10.33594/000000393 |
||||
| 74 Cordaro M, Fusco R, D'Amico R, Siracusa R, Peritore AF, Gugliandolo E, Genovese T, Crupi R, Mandalari G, Cuzzocrea S, Di Paola R, Impellizzeri D: Cashew (Anacardium occidentale L.) Nuts Modulate the Nrf2 and NLRP3 Pathways in Pancreas and Lung after Induction of Acute Pancreatitis by Cerulein. Antioxidants (Basel) 2020;9:992. https://doi.org/10.3390/antiox9100992 |
||||
| 75 D'Amico R, Fusco R, Cordaro M, Siracusa R, Peritore AF, Gugliandolo E, Crupi R, Scuto M, Cuzzocrea S, Di Paola R, Impellizzeri D: Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. Int J Mol Sci 2020;21:2144. https://doi.org/10.3390/ijms21062144 |
||||
| 76 Fusco R, Siracusa R, D'Amico R, Cordaro M, Genovese T, Gugliandolo E, Peritore AF, Crupi R, Di Paola R, Cuzzocrea S, Impellizzeri D: Mucosa-Associated Lymphoid Tissue Lymphoma Translocation 1 Inhibitor as a Novel Therapeutic Tool for Lung Injury. Int J Mol Sci 2020;21:7761. https://doi.org/10.3390/ijms21207761 |
||||
| 77 Peritore AF, D'Amico R, Siracusa R, Cordaro M, Fusco R, Gugliandolo E, Genovese T, Crupi R, Di Paola R, Cuzzocrea S, Impellizzeri D: Management of Acute Lung Injury: Palmitoylethanolamide as a New Approach. Int J Mol Sci 2021;22:5533. https://doi.org/10.3390/ijms22115533 |
||||
| 78 Sawant S, Gokulan R, Dongre H, Vaidya M, Chaukar D, Prabhash K, Ingle A, Joshi S, Dange P, Joshi S, Singh AK, Makani V, Sharma S, Jeyaram A, Kane S, D'Cruz A: Prognostic role of Oct4, CD44 and c-Myc in radio-chemo-resistant oral cancer patients and their tumourigenic potential in immunodeficient mice. Clin Oral Investig 2016;20:43-56. https://doi.org/10.1007/s00784-015-1476-6 |
||||
| 79 Hay J, Shahzeidi S, Laurent G: Mechanisms of bleomycin-induced lung damage. Arch Toxicol 1991;65:81-94. https://doi.org/10.1007/BF02034932 |
||||
| 80 Zhang HY, Gharaee-Kermani M, Zhang K, Karmiol S, Phan SH: Lung fibroblast alpha-smooth muscle actin expression and contractile phenotype in bleomycin-induced pulmonary fibrosis. Am J Pathol 1996;148:527-537. | ||||
| 81 Fusco R, Cordaro M, Siracusa R, Peritore AF, Gugliandolo E, Genovese T, D'Amico R, Crupi R, Smeriglio A, Mandalari G, Impellizzeri D, Cuzzocrea S, Di Paola R: Consumption of Anacardium Occidentale L. (Cashew Nuts) Inhibits Oxidative Stress through Modulation of the Nrf2/HO-1 and NF-kB Pathways. Molecules 2020;25:4426. https://doi.org/10.3390/molecules25194426 |
||||
| 82 Cordaro M, Siracusa R, Fusco R, D'Amico R, Peritore AF, Gugliandolo E, Genovese T, Scuto M, Crupi R, Mandalari G, Cuzzocrea S, Di Paola R, Impellizzeri D: Cashew (Anacardium occidentale L.) Nuts Counteract Oxidative Stress and Inflammation in an Acute Experimental Model of Carrageenan-Induced Paw Edema. Antioxidants (Basel) 2020;9:660. https://doi.org/10.3390/antiox9080660 |
||||
| 83 Rautiainen S, Manson JE, Lichtenstein AH, Sesso HD: Dietary supplements and disease prevention - a global overview. Nat Rev Endocrinol 2016;12:407-420. https://doi.org/10.1038/nrendo.2016.54 |
||||
| 84 Lee SE, Lee HS, Lee YH, Park BS, Choi WS: Antioxidative activities of methanol extracts of tropical and oriental medicinal plants. Applied Biological Chemistry 1998;41:556-559. | ||||
| 85 Hodges RJ, Lim R, Jenkin G, Wallace EM: Amnion epithelial cells as a candidate therapy for acute and chronic lung injury. Stem Cells Int 2012;2012:709763. https://doi.org/10.1155/2012/709763 |
||||
| 86 Johnson KJ, Fantone JC, 3rd, Kaplan J, Ward PA: In vivo damage of rat lungs by oxygen metabolites. J Clin Invest 1981;67:983-993. https://doi.org/10.1172/JCI110149 |
||||
| 87 Fantone JC, Ward PA: Role of oxygen-derived free radicals and metabolites in leukocyte-dependent inflammatory reactions. Am J Pathol 1982;107:395-418. | ||||
| 88 Nguyen T, Sherratt PJ, Huang HC, Yang CS, Pickett CB: Increased protein stability as a mechanism that enhances Nrf2-mediated transcriptional activation of the antioxidant response element. Degradation of Nrf2 by the 26 S proteasome. J Biol Chem 2003;278:4536-4541. https://doi.org/10.1074/jbc.M207293200 |
||||
| 89 Tkachev VO, Menshchikova EB, Zenkov NK: Mechanism of the Nrf2/Keap1/ARE signaling system. Biochemistry (Mosc) 2011;76:407-422. https://doi.org/10.1134/S0006297911040031 |
||||
| 90 Quinlan T, Spivack S, Mossman BT: Regulation of antioxidant enzymes in lung after oxidant injury. Environ Health Perspect 1994;102:79-87. https://doi.org/10.1289/ehp.9410279 |
||||
| 91 Johnson KJ, Ward PA: Acute and progressive lung injury after contact with phorbol myristate acetate. Am J Pathol 1982;107:29-35. | ||||