×
![]()
Corresponding Author: Alicia Ortega-Aguilar
Departamento de Bioquímica, Facultad de Medicina, Universidad Nacional Autónoma de México, Av. Universidad 3000, México City (México)
Tel. , E-Mail aortega@unam.mx
Skeletal Muscle is a Source of α-Synuclein with a Sarcolemmal Non-Lipid Raft Distribution
Ibrahim Ramirez-Soto Alicia Ortega-Aguilar
Departamento de Bioquímica, Facultad de Medicina, Universidad Nacional Autónoma de México, Mexico City, México
Introduction
Alpha synuclein (αSN) is a vertebrate specific protein with no identified invertebrate or prokaryotic homologue [1]. It has received significant scientific attention due its presence in pathological intra and extracellular aggregate structures in Parkinson’s Disease (PD), Lewy Body Dementia, and in Sporadic-Inclusion Body Myositis [2] among other neurodegenerative diseases [3, 4]. When present in the cytoplasm, the 140 aa residues soluble protein has a natively unfolded structure and is comprised of three domains; the N-terminal (aa 1-60) positively charged domain which contains a highly conserved –KTKEGV- motif, known to be involved in membrane binding capacity; the hydrophobic non-amyloid β component region (aa 61-95), and the negatively charged C-terminal (aa 96-140) region, susceptible to post-translational changes such as phosphorylation of serine S129 [5]. Various studies have shown αSN acquires secondary structure in the form of α-helix in the presence of lipids [6-8]. In biological membranes, αSN is present as stable homo-oligomers of different sizes [9, 10]. Although a cytoplasmic protein, αSN is found in a variety of intracellular membranes such as the membranes from endoplasmic reticulum [11, 12], Golgi apparatus [12], mitochondria [12, 13], lysosomes [14], synaptic vesicles [15], nuclei [12, 16] and in plasma membrane isolated from diverse cell cultures and tissues [17-19]. In Addition to its intracellular distribution, αSN has been observed in the extracellular fluids [20], and is secreted to the external environment, like proteins of the 14-3-3 family such as the tau protein, by normal healthy cells [21, 22]. αSN contains sequence homology that relates it with the 14-3-3 protein family which are ubiquitous cytoplasmic chaperones that mediate signal transduction by binding to phosphoserine-containing proteins [23].
A general physiological role of αSN has not yet been clearly defined. Due to its widespread protein interactions and presence in specific signalling pathways, as a monomer it has been associated with multiple physiological functions. In the Central Nervous System (CNS) αSN is synthesized by neurons and glia [24, 25]. In brain (BR) homogenates, a direct interaction between αSN and tyrosine hydroxylase (TH), the rate-limiting enzyme in the synthesis of catecholamines and melanin, has been shown [26]. This interaction inhibits TH activity in a dose-dependent manner thereby regulating the synthesis of dopamine in the substantia nigra pars compacta [27], noradrenaline in the locus coeruleus [28], and adrenaline in the adrenal glands [29]. The physiological consequences of a decrement in catecholamines and melanin synthesis are present in the symptomatology of PD and Lewy Body Dementia [30]. Several other studies show multimeric membrane-bound αSN is a triggering factor in the assembly of the SNARE protein complex, where αSN primarily interacts with Synaptobrevin, a membrane protein present in the storage neurotransmitter vesicle [10]. Interaction of these proteins on the neurotransmitter vesicle leads to binding of the presynaptic membrane, promoting the fusion and release of neurotransmitters. Interestingly, deletion of αSN in mice does not have negative implications for normal CNS activity or survival [31].
Skeletal muscle (SM) accounts for 40% of vertebrate body weight and is therefore the most abundant excitable tissue in the body. The αSN gene, mRNA and protein has been detected in SM [32-34]. In human SM, it has also been detected to be part of the inclusion bodies in the progressive muscle disorder Inclusion-Body Myositis [2]. However, its exact role in muscle function has not been systematically studied. Recently, it has been shown to participate in the traffic of GLUT4 transporter vesicles to the plasma membrane, independent of insulin stimulation [34]. Additionally, when treated with recombinant αSN, SM and pre-adipocytes in culture have increased glucose transport inside the cell through the LPAR2/GAB1/P13K/Akt route, an insulin-independent pathway [35]. In SM and BR, there is a clear involvement of αSN with the plasma membrane.
Ageing is part of normal physiological development of most animals, characterised by deterioration in physiological functions and metabolic processes, as well as an important factor that increases the risk of neurodegenerative diseases in humans. During normal aging, post-mitotic tissues such as SM and BR have been associated with a progressive loss in cell numbers. The progressive effect of aging, mainly on SM is a fundamental feature to understand the frailty observed in old animals.
The association between αSN in SM and BR during normal aging remains to be evaluated, in particular, the distribution and interaction in surface membranes. Understanding the interaction of αSN with lipid membranes and the conformational properties of its bound state in vivo can help identify what promotes transition from physiological to pathological condition during aging in some individuals. In the present study, we investigate in isolated plasma membranes from SM and BR of young and old rats the presence and membrane distribution of αSN to determine whether αSN belongs to a group of proteins that are naturally overexpressed during aging. Furthermore, we studied the contribution of SM towards extracellular αSN that may be of relevance to αSN presence in body fluids.
Materials and Methods
Animals
All procedures were conducted in accordance with the Guide for the Care and Use of Laboratory Animals of the Institute of Laboratory Animal Resources of the United States as approved in Mexico by the Ethics Committee of the School of Medicine of the National Autonomous University of Mexico (UNAM) (NOM-062-ZOO1999).
Male Wistar rats 3 and 24 months old, considered to be young and old, were euthanized by cervical dislocation and fast skeletal muscle and brain were isolated at room temperature.
Isolation of Skeletal Muscle membrane fractions
Isolation of Skeletal Muscle Cytosol and Sarcolemma. The Sarcolemma was obtained from fast skeletal muscle (forelimbs, hind limbs, and back muscles) by differential centrifugation and a discontinuous sucrose gradient as previously described [36]. Briefly, for one isolation, cleaned muscle obtained from 4 rats were homogenized with a polytron for 10 seconds in a solution containing in mM: 20 Tris-malate and 100 KCl, pH 7.0., followed by a 10 second rest; this procedure was repeated two times. The homogenate was centrifuged at 12,000 × g for 20 min at 4°C. The supernatant was collected and filtered through four layers of gauze. The filtered supernatant was then centrifuged at 140,000 × g for 40 min at 4°C. Some of the supernatant was retrieved and stored as skeletal muscle cytosol (CYT) and tested to be devoid of Flotillin-2.
Solid KCl was added to a final concentration of 0.6 M to the remaining supernatant and the sample was incubated for 1 h with continuous stirring on ice. The homogenate was then centrifuged at 140,000 × g for 40 min at 4°C. The precipitated fraction was suspended in a solution containing in mM: 20 Tris-malate and 100 KCl, pH 7.0 (Buffer A), to eliminate excess KCl. the collected precipitated fraction was suspended in a solution containing 20 mM Tris-malate and 250 mM sucrose, pH 7.0, transferred to a sucrose gradient of 23%, 26%, 29% and 35% w/v and centrifuged at 75,000 g for 16 h at 4°C. The 23/26% interphase has been proven to correspond to the Sarcolemma fraction (SL), enriched in β-dystroglycan and low dihydropyridine receptor content as determined by [36]. Each fraction in the gradient was collected separately and suspended in Buffer A and centrifuged at 140 000 × g for 40 min at 4°C. The precipitated membranes from each interphase were collected and suspended in 500 µL of Buffer A, and stored at -20°C to further test for purification.
The protein concentration was determined with the Coomassie Plus Protein Assay Reagent (Pierce, Rockford, IL, USA) with BSA as the standard.
Isolation of transverse tubules membranes. Microsomes of SM homogenate were obtained by differential centrifugation as described in section 2.2.1, and placed on a discontinuous sucrose gradient of 25, 27.5, 30, and 35% (w/v) and centrifuged at 75,000 x g for 16 h. The membranes isolated from the sucrose gradient at the 25/27.5% interphase corresponded to the transverse tubule (TT) as previously characterized in rat SM [37].
Isolation of Brain Cytosol, Myelin and myelin-free brain cell membranes (BNm)
Whole brain plasma membranes and CYT were isolated using a modified protocol [38] first described by [39]. Briefly, male Wistar rats weighing 240 to 280 g were euthanized by cervical dislocation. For one isolation, fresh or frozen brains obtained from 4 rats were first minced and then homogenized with a glass-Teflon homogenizer by hand with 20 strokes in 15 mM Tris-HCl, 320 mM sucrose buffer, pH 7.8 and centrifuged three times to discard nuclei and other debris at 3000 x g for 10 min at 4°C. A fraction of the collected supernatants was combined and centrifuged at 140,000 × g for 40 minutes at 4 °C to obtain CYT. The supernatant was retrieved and tested to be devoid of Flotillin-2 and stored as brain CYT.
Remaining supernatants were layered over an 850 mM, Tris-HCl 15mM, pH 7.8 sucrose cushion and centrifuged for 1 h, 70,000 x g, at 4°C. The interphase cloud between 320 mM and 850 mM sucrose buffers corresponds to myelin which has been proven to be enriched in two of the main myelin protein markers: myelin binding protein and proteolipid and was stored [38]. To eliminate mitochondria and possible residual myelin for further myelin-free brain plasma membrane (BNm), the remaining sample was centrifuged for 30 min at 20,000 x g, 4 °C. The supernatant was recovered and centrifuged at 100,000 × g, 4 °C, for 1 h to sediment the BNm.
Isolation of detergent-resistant membrane (DRM)
DRM were isolated as previously described [40]. Briefly, aliquots of 0.1 mg/ml isolated SL or BNm were incubated for 30 min with a solution containing 1% Triton X-100 on ice. After incubation, the membranes were gently mixed with an equal volume of 80% sucrose (w/v) to give a final sucrose concentration of 40% and placed at the bottom of an ultracentrifuge tube. The membranes were overlaid with 2.5 ml 30 % sucrose, followed by 1 ml 5 % sucrose and centrifugation at 200,000 ×g for 18 hr. Nine 0.5-ml fractions (excluding the pellet) were collected from the top of the gradient where fraction 1 corresponds to the top of the tube and fraction 9 the bottom. The protein was suspended in an equal volume for each fraction, and the protein content was analysed by SDS-PAGE and immunoblotting under reducing conditions.
Plasma membrane cholesterol depletion
To study the effect cholesterol removal has on αSN membrane distribution, isolated SL or BNm was incubated for 1 h at 37 °C with 30 mM Methyl-β-cyclodextrin (MβCD, Sigma-Aldrich) with agitation. The membrane was then washed with a PBS buffer, pH 7.0, centrifuged at 100,000 × g, 4 °C for 1 h, and underwent the DRM isolation protocol as described in section 2.4.
Immunoblots and densitometry
Fraction samples were separated by 10% SDS-PAGE on a 10% polyacrylamide gel and transferred to a nitrocellulose membrane. After transfer, nitrocellulose membranes were either used directly or fixed with 0.5% paraformaldehyde in PBS buffer and incubated for 30 min (in any case there was no difference and finally used without paraformaldehyde). Membranes or recombinant αSN protein (abcam 51189) for Western blot or dot blot were blocked for at least 40 minutes with 5% nonfat dry milk (Bio-Rad) and incubated with the corresponding antibody or toxin. Anti- αSN primary antibody (1:500, 2B2D1, sc-53955, monoclonal, Santa Cruz), Flotillin-2 (1:1000, B-6, sc-28320, monoclonal, Santa Cruz), anti-αSN (Phospho-S129) (1:1000, ab-51253, monoclonal, abcam), Transferrin receptor (1:1000, H68.4, ab-269513, monoclonal, abcam), GAPDH antibody [GT239] Gene Tex (1:1000), Peroxidase conjugated cholera toxin B subunit (1:50,000, C3741, Sigma Aldrich). After washing and incubation with the corresponding peroxidase-labelled secondary antibody AffiniPure Goat Anti-Mouse (115-035-003, Jackson Immunoresearch), blots were developed using the C-DiGit Blot scanner (LI-COR Biosciences). Densitometry analysis of Western blots were performed by using the Image studio software (Li-COR Biosciences). The same amount of area was defined as antibody response for each of the different blots and used the same area for bands located on the same Western blot. Densitometry values (O.D.) of bands were corrected for background by subtracting the non-reactive part of the membrane for that blot.
All SDS-PAGE were loaded using the same protein concentration. Equal protein loading was confirmed by Ponceau-S staining of the membranes. The O.D. values for every protocol used are expressed as means ± standard deviation (SD).
Intact muscle experiments
Isolation of Extensor digitorum longus muscle. Male Wistar rats weighing 280–300 g of 3-months old and male Wistar rats weighing 550–600 g of 24-months old, were euthanized by cervical dislocation, and the EDL muscles were isolated at room temperature. The isolated muscle was placed into an acrylic chamber that was equipped with platinum electrodes along each side of the chamber wall to allow contact with the Krebs solution as previously described [41]. Briefly, we used Krebs solution containing in mM: 135 NaCl, 5 KCl, 1 MgCl2, 2.5 CaCl2, 11 dextrose, 1 NaPO4 dibasic, and 15 NaHCO3 and a gas mixture of 95% O2 and 5% CO2 to reach a pH of 7.0. Isolated EDL muscle was fastened by its distal tendon to forceps and by its proximal tendon to a force transducer (FT-03, Grass Medical Instruments, RI, USA). The platinum electrodes were connected in parallel to two stimulators (S88, Grass Medical Instruments, RI, USA).
EDL muscle incubation medium to detect αSN. EDL muscle placed in the experimental chamber was kept in Krebs solution, the incubation medium was collected before the first stimulation protocol and after 3 h incubation and stored the incubation solution as extracellular media isolated from skeletal muscle. EDL muscle from young and old rats were examined for viability by mechanical stimulation time zero and after the 3 h incubation. The incubation medium was used to detect α-SN by Western blot analysis.
EDL muscle stimulation protocol. Single twitch (ST) of 0.6 ms were used to reach the voltage for maximal tension. To obtain the optimal sarcomere length (2.4 μm), the muscles were stretched to the length at which the twitch force was maximal. The muscles were stretched to the optimum length at which the three ST force was maximal (1 Hz and 100 V), followed by tetanic stimulations (T1 to T5) of 75 Hz for 3 s at 90 V followed by 2 min rest. At the end of the protocol, the muscle was rested for 10 min before new stimulation was applied to probe for muscle force recovery after fatigue (R).
All values for every protocol used are expressed as means ± standard deviation.
Results
Presence of αSN in Skeletal muscle and Brain cytosol and plasma membrane
In Fig. 1A, to confirm whether the separation of the CYT and membranes in BR and SM was performed accurately, we evaluated by western blot the membrane marker protein flotillin which was absent in the CYT of SM and BR, meaning that there was no membranes contamination, whereas its presence is detected in the isolated SL and BNm. Fig. 1B shows a representative western blot and the statistical representation of αSN content in CYT and SL. In SL, αSN signal was two times more when compared to the αSN in the CYT with equal mg of protein is loaded. In SM αSN is detected as a monomer and oligomers mainly tetramer (TM) and pentamer (PM), where the PM structure is mainly found in SL and the monomer in CYT. The recombinant αSN was used as standard to confirm αSN recognition. Considering the presence of αSN in BR from normal rats, Fig. 1C shows a representative western blot and the statistical representation of αSN content in CYT and BNm. In BNm, αSN is present in similar proportions to the CYT with equal mg of protein loaded. However, in BR CYT, αSN exists as a monomer and TM. In BNm αSN is detected as TM and PM. Because αSN can exist as a phosphorylated protein (P-αSN), we investigated whether P-αSN exists in normal SM and BR CYT and membranes using the antibody against P-αSN129 (EP1536Y).
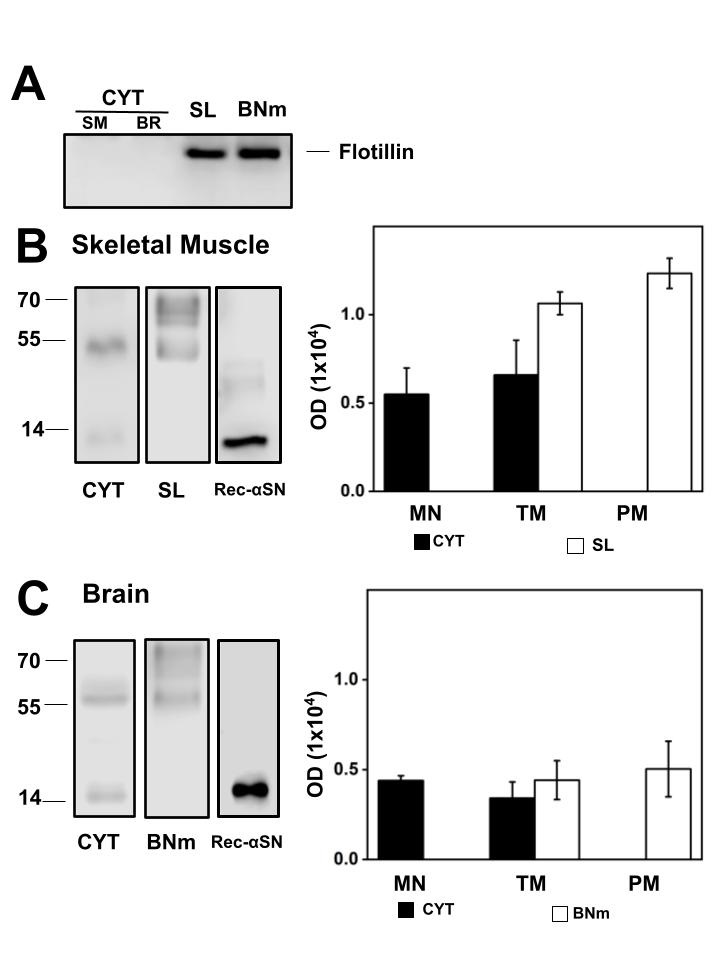
Presence of P-αSN in Skeletal muscle and Brain cytosol and plasma membrane
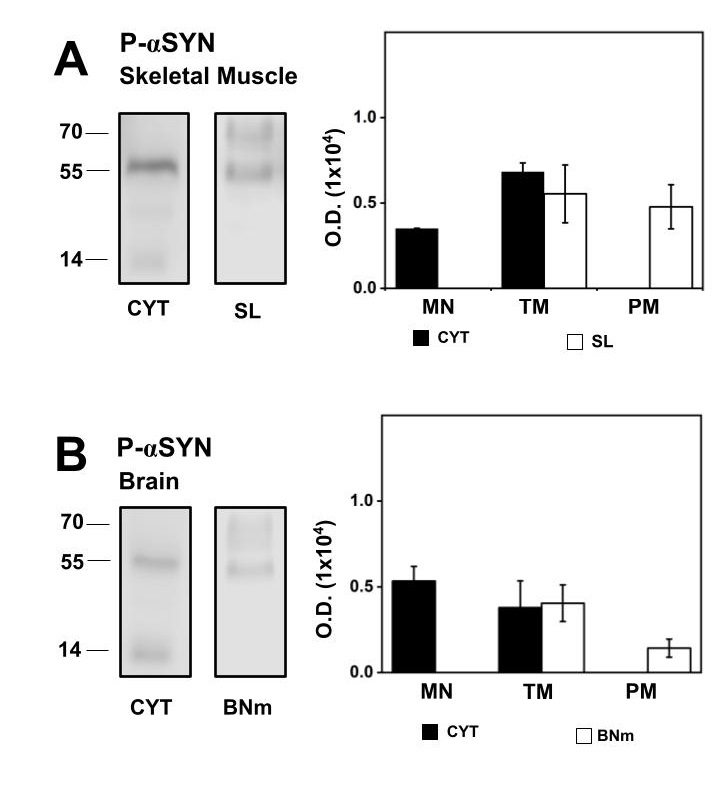
Fig. 2A shows a representative western blot and the statistical representation of P-αSN content in CYT and SL. In CYT, the monomer and TM structure of P-αSN were present, while TM and PM structures were present in SL. The amount of total P-αSN in CYT was similar to the total amount of P-αSN observed in SL, equal protein loaded. Contribution of each individual structural form of P-αSN was calculated; the P-αSN monomer comprises 34%, while the P-αSN TM corresponds to the remaining 66% of P-αSN in CYT. In SL the P-αSN TM comprises 53%, while the P-αSN PM corresponds to the remaining 47% of P-αSN detected.
In BR, Fig. 2B corresponds to a representative western blot and the statistical representation of P-αSN content in CYT and BNm with equal protein loaded. In brain P-αSN monomer was mainly present in CYT which accounts for 58%, while the TM P-αSN corresponds to the remaining 42%. In BNm the P-αSN TM comprises 84%, while the P-αSN PM corresponds to the remaining 16%. Thus, our results show that αSN, along with the P-αSN, were present in CYT, SL and BNm in normal rats. Since αSN oligomers were a part of an isolated membrane of healthy rats, we determined if αSN remains present in isolated plasma membranes from old rats.
Comparative αSN expression in isolated myelin-free brain cells plasma membranes and isolated Myelin
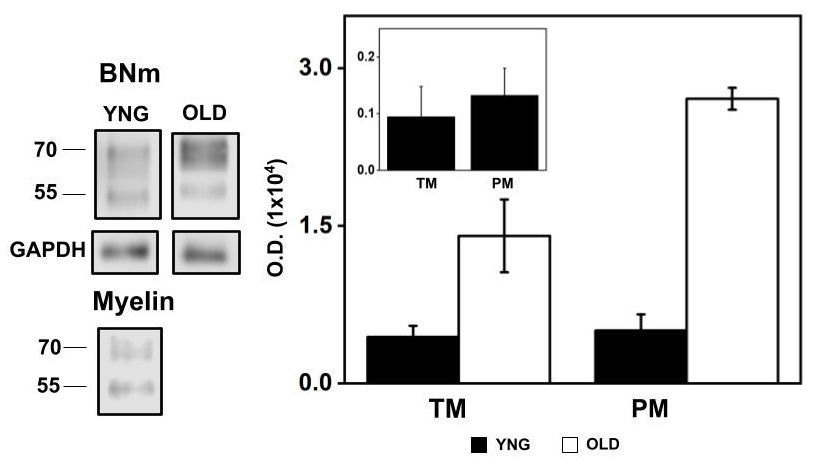
Fig. 4 shows a representative western blot of αSN expressed in BNm and Myelin and the statistical representation of the immunoblot density in young and old BNm, by equal mg protein loaded. Inset graph corresponds to the statistical representation of the αSN immunoblot density of Myelin obtained from young rat equal mg protein loaded. In BNm, there was determined a similar amount of αSN TM and PM in young rat whereas in the old rat αSN PM was double the expression with respect to the TM. Inset plot shows αSN expression in Myelin was five-fold lower in both TM and PM structures when compared to BNm from young rats as determined by equal mg protein loaded.
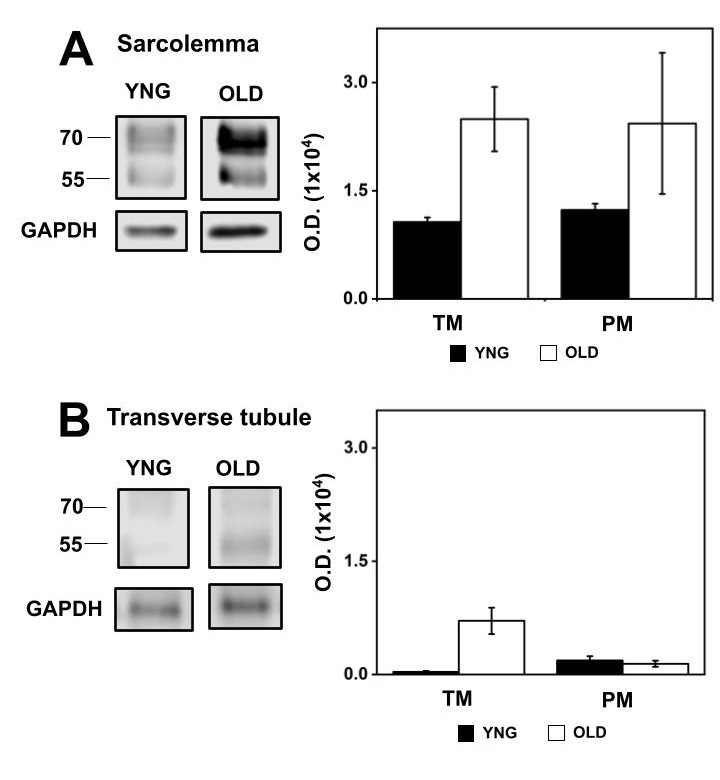
By separating natively high cholesterol content plasma membranes such as TT-membranes in SM and Myelin in BR we demonstrated that αSN is poorly expressed in high cholesterol content membranes. To determine the distribution of αSN in muscle and neural cells, DRM from SL and BNm were isolated and characterized.
Distribution of αSN in Sarcolemma and myelin-free brain cells plasma membrane treated with Triton X-100
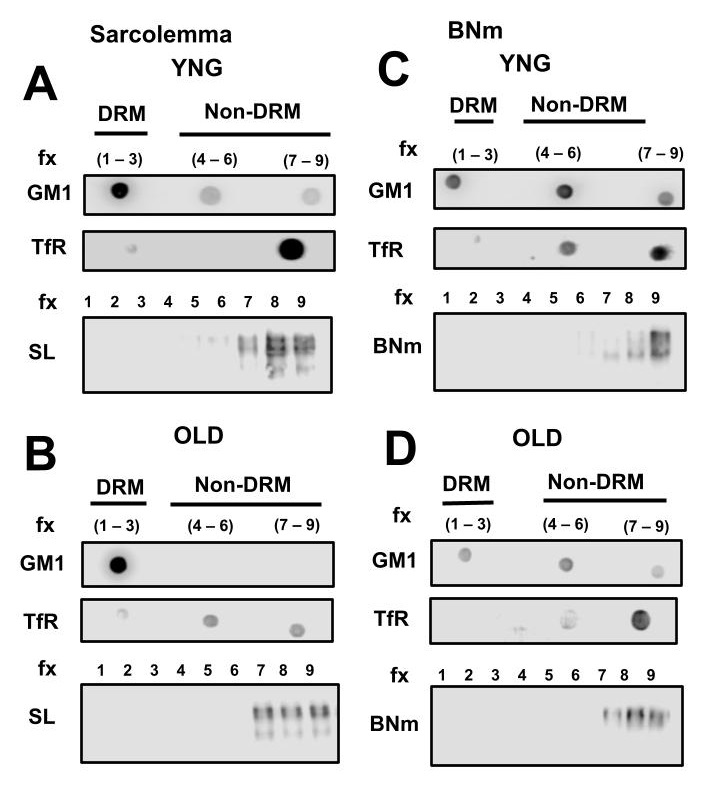
Fig. 5 shows αSN distribution solely in the non-detergent resistant membrane (non-lipid raft) fractions isolated from SL of young (Fig. 5A) and old (Fig. 5B) SM and from BNm of young (Fig. 5C) and old (Fig. 5D) brains. In all cases, dot blot analysis of ganglioside GM1, a lipid raft marker, was detected in the fractions 1-3 and the transferrin receptor (TfR), a non-lipid raft protein marker was distributed mainly in the fractions 7-9 in SL and BNm. To better visualize the distribution of αSN along the 1-9 fraction gradient, western blot analysis of αSN shows its distribution mainly localized in the 7-9 fractions for SL (Fig. 5A) and BNm (Fig. 5C). As observed, αSN was a non-raft protein in both SL and BNm. With respect to αSN distribution during aging, western blot analysis of αSN showed it remains in the 7-9 fractions for old SL (Fig. 5B) and old BNm (Fig. 5D). To prove cholesterol concentration has an effect, if any, on αSN membrane distribution, we depleted cholesterol from the membrane.
Distribution of αSN in Sarcolemma and myelin-free neural cell plasma membrane from young rats incubated with MβCD and treated with Triton X-100
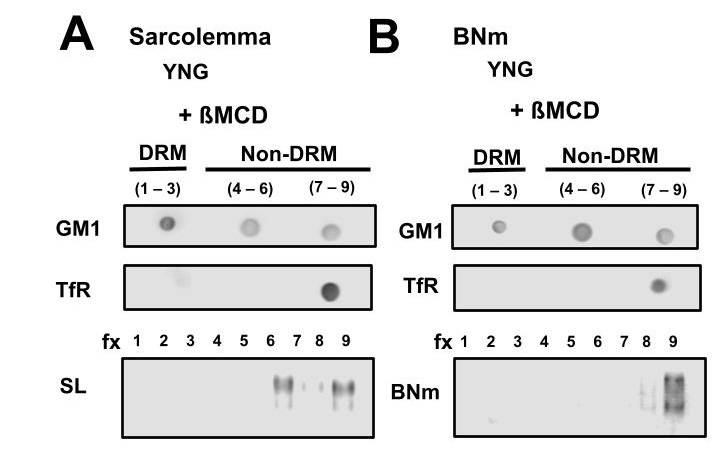
Fig. 6 shows αSN was distributed in the non-raft fractions of SL (Fig. 6A) and BNm (Fig. 6B) after incubation with 30 mM MβCD. Dot blot analysis of GM1 and TfR was detectable in the fractions 1-3 and 7-9 respectively in SL and BNm. Interestingly in both membranes, there was no movement of αSN towards the detergent resistant membrane (lipid raft).
Extracellular αSN in incubation medium from intact functional skeletal muscle from old rats
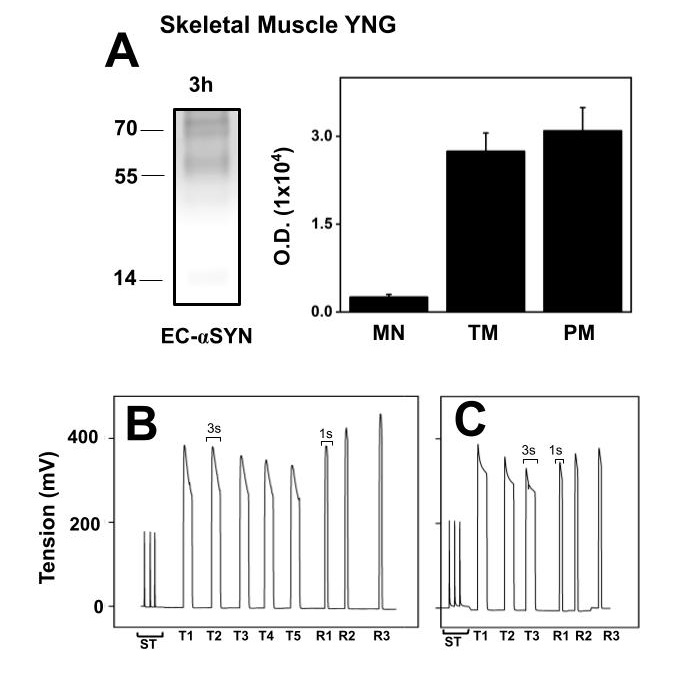
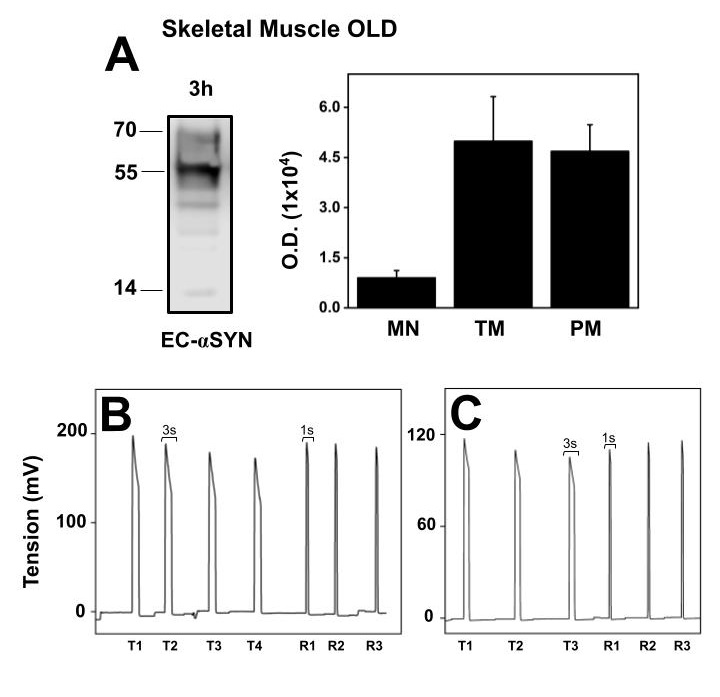
Since αSN levels increase old SM, we determined if in isolated muscle under incubation conditions in Krebs solution, αSN released to the extracellular medium varies with age. Fig. 8A shows representative western blot and the statistical representation of the αSN structures present in extracellular media obtained from intact old SM, specifically Extensor digitorum longus (EDL) muscle. The major αSN structures detected were TM and PM. When compared to extracellular monomer αSN, the oligomers were twelve times more abundant. Fig. 8B is the registered tension of EDL mechanical parameters obtained before the incubation period, shows maximal tension, fatigue and fatigue recovery of a freshly dissected EDL muscle. The maximal tension observed is half the maximal tension observed in young EDL. Fig. 8C shows the stimulation of muscle after the 3 h incubation to test the functionality of the EDL muscle. Although tension decreased in average 30%, we observed that after the incubation there was a full recovery after fatigue, indicated that EDL muscle was partially affected after three-hour incubation. αSN present in the incubation medium is not the result of muscle damage in the old muscle.
Distribution of α-SN into non-detergent resistant membranes from Sarcolemma and myelin-free brain plasma membranes
In BR and SM from young and old rats, we find α-SN partitions in the non-DRM (non-lipid rafts). In SM, TT-membranes behave like lipid rafts, since these membranes are enriched in cholesterol, sphingomyelin and caveolin-3. As described in the previous paragraph, TT-membranes isolated from healthy young and old rats show poor expression of αSN. It has also been observed that Myelin which contains the largest amount of cholesterol in the brain has a reduced expression of αSN. To the contrary of what we have observed, it has been reported that αSN is a lipid raft located protein detected in DRM from two different cell types; neurons isolated from mice and HeLa cells, incubated with recombinant αSN [61]. It is important to review the images that demonstrate α-SN is a DRM protein in neurons which co-localized with lipid raft markers [61]. Here, it is possible to observe that αSN is distributed mainly along the non-DRM. Therefore, the conclusion that αSN is located exclusively in lipid rafts is not an accurate interpretation of the result. Additionally, the αSN detected in the experiments are not endogenous to the cell, the mice used to isolate brain cell membranes are transgenic mice overexpressing wild type human αSN. It is also possible that the amount of αSN in the membranes does not reflect what exists in a normal cell within tissues and this fact may alter its distribution. By using a similar extraction protocol as in the present study, detergent treated membrane fractions were obtained from HeLa cells transfected with αSN to observe its distribution in the plasma membrane. A similar result is observed where there is presence of αSN in most of the fractions including non-DRM. In fact, most of αSN partitions into the non-DRM seen by band optical density and size, consequently, α-SN is not localized in lipid rafts exclusively as described [61]. The presence of αSN in the DRM fraction seen in this study can be due to a similar effect as seen in the case of transfected mice. Thus, the data and interpretation claiming αSN is a lipid raft protein is not accurate in their experimental conditions, furthermore it is not indicative of what may be occurring under physiological tissue conditions. Our results do share the same result, using the same isolation protocol, in non-DRM obtained from plasma membrane isolated from healthy skeletal muscle and brain, αSN partitions in the non-DRM co-localizing with the TfR, a classical non-lipid raft marker.
In vivo studies suggest that αSN can exist embedded into the hydrophobic core of the lipid membrane. Membrane fractions isolated from the rat brain were used to test the involvement of electrostatic force in the interaction between αSN and a biological membrane [62]. To do this, membrane fractions were incubated in different salt concentrations, and were then isolated and probed for remaining αSN. This experiment showed that αSN remained bound to membranes throughout the ionic strength range tested [62]. If αSN were a peripheral membrane protein, the expected result would be αSN release in the supernatant. This salt-wash experiment confirms αSN interaction with membranes is of a hydrophobic nature, since hydrophobic interactions are not interrupted by changes in the ionic strength of the solvent. In the isolation protocol of SL and TT-membrane, homogenized SM is incubated with 0.6 M KCl to extract and eliminate the major contractile proteins. Under these conditions, any membrane associated protein that is bound by electrostatic forces would be displaced from the membrane into the solvent [63]. Our results also indicate that αSN interaction with biological membranes is mainly through hydrophobic interactions as can be observed for other brain cell plasma membrane proteins [36]. In addition, molecular dynamic simulations reveal that αSN can penetrate the cell membrane rapidly. Within 4ns of simulation αSN can penetrate up to 2/3 of the membrane and within 9 ns can fully penetrate the lipid bilayer [64]. The molecular dynamic model of αSN insertion into lipid bilayer reveals that membrane penetration promotes the incorporation of additional αSN monomers into the complex [64]. These studies provide a molecular explanation for the incorporation and multimerization of αSN in a lipid membrane. In vitro studies with α-SN reveal the trypsin digestion with small unilamellar vesicles composed of phosphatidylcholine and phosphatidylserine results in a population of partially degraded and whole αSN is observable even after 30 minutes of trypsin digestion suggesting two things: A population of αSN remains embedded in the membrane and another population of αSN can cross lipid membranes protein and located inside, protected from protease activity [65].
Effect of cholesterol depletion on Sarcolemma and myelin-free brain plasma membranes with MβCD does not modify αSN membrane distribution
The surface-acting molecule MβCD can selectively extract membrane cholesterol by incorporating it in a nonpolar cavity of cyclic oligomers of glucopyranoside 1,4 glycosidic binding [66]. Sarcolemma cholesterol depletion with 30 mM MβCD has been previously shown to reduce the cholesterol content by 12% with an important effect on membrane protein distribution related to lipid raft [36]. The same 30 mM MβCD in BNm reduced cholesterol content by 80% [38]. The MβCD-resistant cholesterol in total plasma membrane or in Triton X-100-isolated DRMs from isolated membranes, represents the cholesterol tightly associated with sphingolipids forming the lipid rafts, whereas the rest of the plasma membrane cholesterol is readily extracted by MβCD [67]. We demonstrate in this study that depletion of cholesterol in SL and BNm, which affects lipid raft proteins distribution, [68] does not affect αSN content and distribution outside the DRM, indicating that cholesterol modification in the DRM does not have a substantial change to allow αSN distribution in the lipid raft membrane fractions, these characteristics is also shared by TfR. Because of the fact αSN is not a lipid raft protein, α-SN is hypothetically excluded from participating in intracellular signalling [69].
Extracellular αSN is detected in the incubation medium from intact functional skeletal muscle
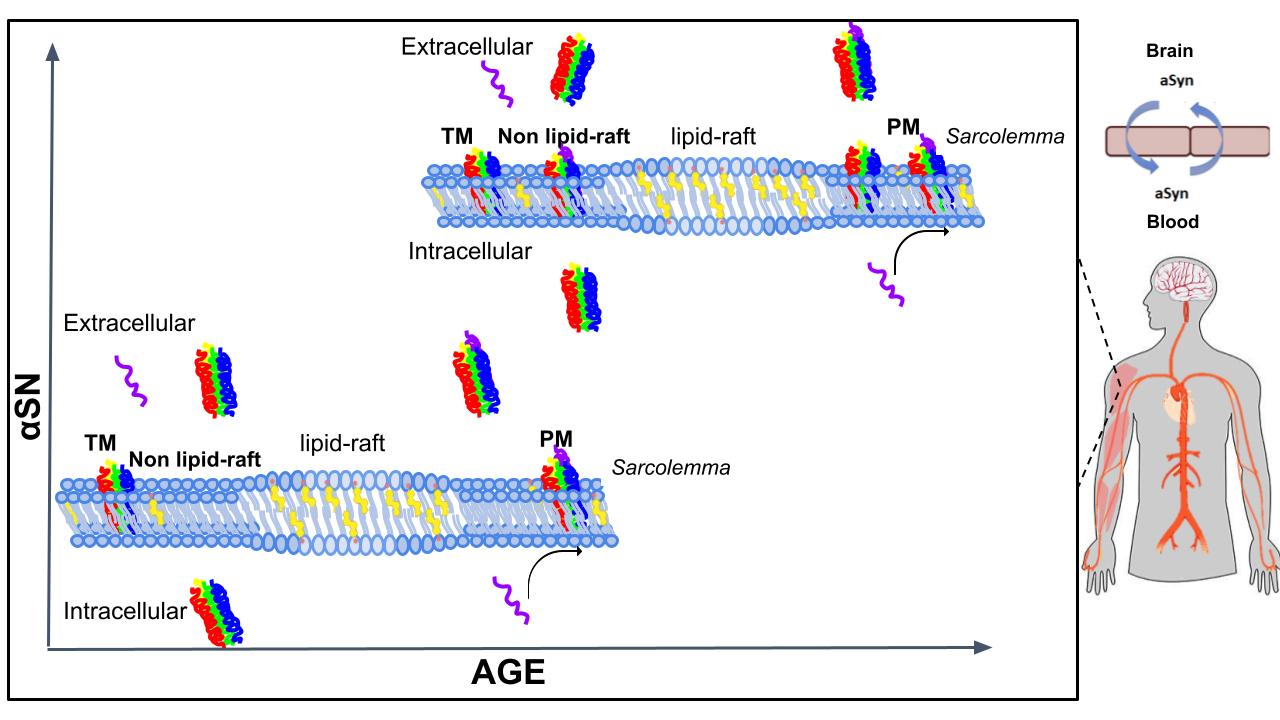
Throughout the years, several studies demonstrate that when α-SN is overexpressed in cells, there is oligomerization of α-SN, leading to proteinaceous aggregates in the cytoplasm and ultimately cell death [70, 71]. At the membrane level, other studies demonstrate in vivo and in silico that αSN forms oligomeric associations in the membrane, forming a pore-like structure that disrupts the ion homeostasis in cells leading to cytotoxicity [64]. Increasing studies seem to suggest that the oligomeric association of αSN is the initiating factor in the development of degenerative diseases, acting as a seed that promotes the misfolding of αSN into pathological fibre structures [72]. In the results presented in this study we show that intact functional isolated EDL muscle incubated in Krebs solution after 3 h, shows an important extrusion of α-SN as monomer, TM and PM structures which is higher in old EDL muscle. The mechanical activity of the EDL muscle does not have a significant change in force, and recovery from fatigue after the incubation protocol, meaning that the soluble α-SN found in the Krebs solution was not due to muscle damage or death. The existence of α-SN as extracellular multimeric complexes implies that these associations can cross SL through the non-raft domains. Translocation of α-SN across the plasma membrane of several cell systems has been observed previously [73]. The study shows that fluorescently tagged αSN is transported internally into a variety of cells, including HeLa, neuronal, hematopoietic, and Chinese hamster ovary cells [73]. A confocal analysis of the fluorescently tagged αSN revealed that αSN begins to appear inside cells within 5 minutes of incubation. Flow cytometric analysis of control cells and incubated with αSN incubation corroborates the confocal microscopic observations. Since the translocation of α-SN into cells is not cell type specific this suggests that αSN does not require a specific cell receptor for transport into cells. Interestingly, the same study discovered that translocation into cells requires one or more of the repeat sequence motifs located in the amino terminal of αSN [73]. The uptake of αSN into cells was independent of receptor-mediated endocytosis. These features of αSN prove that it is behaving as a cell penetrating protein. Combining this fact with other Information provided by the transport of αSN, the SM as a source of extracellular αSN raises an interesting concern specially during aging. The relevance of aging muscle is implicated in the appearance of αSN in the extracellular environment, which become incorporated into biological fluids, eventually reaching the blood-brain barrier. This new information is a fact that may be worth considering further investigation in neurodegenerative diseases in the elderly. It is established that αSN crosses the blood-brain barrier in the blood-to-brain direction [74]. Even though the exact mechanism for αSN to cross the SL is unknown, the transport of αSN across cell membranes is possible [73]. The SL lipid composition with age changes to produce a more rigid and ordered plasma membrane [75], resembling the ordered stable structure of lipid rafts. However, aSN was found outside the lipid raft meaning that the change in fluidity does not affect αSN membrane distribution, rather the cholesterol/sphingolipid content. Given this information, it is possible that the biophysical properties in SL, which may not necessarily be related to cholesterol concentration change with age and may promote the dissociation of αSN from SL to the extracellular medium. For a schematic representation of our findings and potential implications in human pathophysiology, see Fig. 9.
Conclusion
The equilibrium that normally exists between membrane-associated and membrane-free α-SN during younger stages in life is shifted towards membrane-free during ageing. Depletion of membrane cholesterol of excitable cells may interfere with αSN structure formation and release. This could potentially increase the concentration of extracellular αSN, which could be very significant considering the amount of muscle mass that exists in vertebrates.
Abbreviations
αSN (Alpha synuclein); BNm (Myelin-free brain cell membrane); BR (Brain); CNS (Central nervous system); CYT (Cytosol); DRM (Detergent resistant membrane); EDL (Extensor digitorum longus ); MβCD (Methyl-β-cyclodextrin); MN (Monomer); P-αSN (Phosphorylated αSN); PD (Parkinson’s disease); PM (Pentamer); SL (Sarcolemma); SM (Skeletal muscle); TfR (Transferrin receptor); TH (Tyrosine hydroxylase); TM (Tetramer); TT (Transverse tubule).
We thank Dr. Aura Jiménez-Garduño from the Universidad de las Américas, Puebla, Mexico and Dr. Fidel Ramón y Romero for fruitful discussion, to Dr. Isabel Arrieta-Cruz from the Instituto Nacional de Geriatria de México who partially contributes with brain tissue from old rats,.
Author Contributions
I.R.S. Conceptualization, methodology, formal analysis, data curation, Writing-Review and Editing. A.O.A. Conceptualization, methodology, formal analysis, Writing-Review and Editing, Validation, Supervision, Project administration.
Funding
This study was supported by Dirección General de Asuntos del Personal Académico, grant DGAPA-IN219119 (AO), Universidad Nacional Autónoma de México (UNAM), the Thematic Network on Proteins, Prions and Neurodegenerative Diseases, directed by Dr. José Antonio de la Peña Mena, Consejo Nacional de Ciencia y Tecnología (PRyEND-CONACyT-280087) network member (AO); Ibrahim Ramirez-Soto was supported during his Master Degree (Graduate Studies in Biochemical Sciences UNAM) by scholarships from the CONACyT, (CVU 968876) and DGAPA-IN219119.
The authors declare that no conflicts of interest exist.
| 1 Yuan J, Zhao Y: Evolutionary aspects of the synuclein super-family and sub-families based on large-scale phylogenetic and group-discrimination analysis. Biochem Biophys Res Commun 2013;15:308-317. https://doi.org/10.1016/j.bbrc.2013.09.132 |
||||
| 2 Askanas V, Engel K, Alvarez R, McFerrin J, Broccolini A: Novel Immunolocalization of α-Synuclein in Human Muscle of Inclusion-Body Myositis, Regenerating and Necrotic Muscle Fibers, and at Neuromuscular Junctions. J Neuropathol Exp Neurol 2000;59:592-598. https://doi.org/10.1093/jnen/59.7.592 |
||||
| 3 Yoshimoto M A, Kang ID, Otero D, Xia Y, Saitoh T: NACP, the precursor protein of the non-amyloid beta/A4 protein (A beta) component of Alzheimer disease amyloid, binds A beta and stimulates A beta aggregation. Proc Natl Acad Sci U S A 1995;92:9141-9145. https://doi.org/10.1073/pnas.92.20.9141 |
||||
| 4 Spillantini M, Crowther RA, Jakes R, Cairns NJ, Lantos PL, Goedert M: Filamentous α-Synuclein Inclusions Link Multiple System Atrophy with Parkinson's Disease and Dementia with Lewy Bodies. Neurosci Lett 1998; 251:205-208. https://doi.org/10.1016/S0304-3940(98)00504-7 |
||||
| 5 Emanuele M, Chieregatti E: Mechanisms of alpha-synuclein action on neurotransmission: cell-autonomous and non-cell autonomous role. Biomolecules 2015;5 :865-892. https://doi.org/10.3390/biom5020865 |
||||
| 6 Galvagnion C, Buell AK, Meisl G, Michaels TC, Vendruscolo M, Knowles TPJ, Dobson CM: Lipid vesicles trigger αsynuclein aggregation by stimulating primary nucleation. Nat Chem Biol 2015;11:229-234. https://doi.org/10.1038/nchembio.1750 |
||||
| 7 Bartels T, Ahlstrom LS, Leftin A, Kamp F, Haass C, Brown MF, Beyer K: The N-terminus of the intrinsically disordered protein α-synuclein triggers membrane binding and Helix folding. Biophys J 2010;99;2116-2124. https://doi.org/10.1016/j.bpj.2010.06.035 |
||||
| 8 Bartolus M, Tombolato F, Tessari I, Bisaglia M, Mammi S, Bubacco L, Ferrarini A, Maniero AL: Broken Helix in Vesicle and Micelle-Bound α-Synuclein: Insights from Site-Directed Spin Labeling-EPR Experiments and MD Simulations. J Am Chem Soc 2008;6690-6691. https://doi.org/10.1021/ja8010429 |
||||
| 9 Kim S, Yun SP, Lee S, Umanah GE, Bandaru V, Yin X, Rhee P, Karuppagounder SS, Kwon SH, Lee H, Mao X, Kim D, Pandey A, Lee G, Dawson VL, Dawson TM, Ko SH: GBA1 deficiency negatively affects physiological α-synuclein tetramers and related multimers. Proc Natl Acad Sci USA 2018;115:798-803. https://doi.org/10.1073/pnas.1700465115 |
||||
| 10 Burré J, Sharma M, Südhof TC: α-synuclein assembles into higher-order multimers upon membrane binding to promote SNARE complex formation. Proc Natl Acad Sci U S A 2014;11:E4274-E4283. https://doi.org/10.1073/pnas.1416598111 |
||||
| 11 Oaks AW, Marsh-Armstrong N, Jones JM, Credle JJ, Sidhu A: Synucleins antagonize endoplasmic reticulum function to modulate dopamine transporter trafficking. PLoS One 2013;8:e70872. https://doi.org/10.1371/journal.pone.0070872 |
||||
| 12 Mori F, Inenaga C, Yoshimoto M, Umezu H, Tanka R, Takahashi H, Wakabayashi K: α-Synuclein immunoreactivity in normal and neoplastic Schwann cells. Acta Neuropathol 2002;103:145-151. https://doi.org/10.1007/s004010100443 |
||||
| 13 Devi L, Raghavendran V, Prabhu BM, Avadhani NG, Anandatheerthavarada HK: Mitochondrial Import and Accumulation of a-Synuclein Impair Complex I in Human Dopaminergic Neuronal Cultures and Parkinson Disease Brain. J Biol Chem 2008;9089-9100. https://doi.org/10.1074/jbc.M710012200 |
||||
| 14 Senol AD, Samarani M, Syan S, Guardia CM, Nonaka T, Liv N, Latour-Lambert P, Hasegawa M, Klumperman J, Bonifacino JS, Zurzolo C: α-Synuclein fibrils subvert lysosome structure and function for the propagation of protein misfolding between cells through tunneling nanotubes. PLoS Biol 2021;19:e3001287. https://doi.org/10.1371/journal.pbio.3001287 |
||||
| 15 Burré J, Sharma M, Tsetsenis T, Buchman V, Etherton MR, Südhof TC: Alpha-synuclein promotes SNARE-complex assembly in vivo and in vitro . Science 2010;329:1663-1667. https://doi.org/10.1126/science.1195227 |
||||
| 16 Maroteaux L, Campanelli JT, Scheller RH: Synuclein: A Neuron-Specific Protein Localized to the Nucleus and Presynaptic Nerve Terminal. J Neurosci 1988;8:2804-2815. https://doi.org/10.1523/JNEUROSCI.08-08-02804.1988 |
||||
| 17 Jacob RS, Eichmann C, Dema A, Mercadante D, Selenko P: α-Synuclein plasma membrane localization correlates with cellular phosphatidylinositol polyphosphate levels. Elife 2021;10:e61951. https://doi.org/10.7554/eLife.61951 |
||||
| 18 Li R, Feng HJ, Federoff SV, Maguire-Zeiss KA, Howard VA: α-Synuclein mediates alterations in membrane conductance: a potential role for α-synuclein oligomers in cell vulnerability. Eur J Neurosci 2010;32:10-17. https://doi.org/10.1111/j.1460-9568.2010.07266.x |
||||
| 19 Volpicelli-Daley LA, Gamble KL, Schultheiss CE, Riddle DM, West AB, Lee VM: Formation of α-synuclein Lewy neurite-like aggregates in axons impedes the transport of distinct endosomes. Mol Biol Cell 2014;25:4010-4023. https://doi.org/10.1091/mbc.e14-02-0741 |
||||
| 20 El-Agnaf OMA, Salem SA, Paleologou KE, Cooper LJ, Fullwood NJ, Gibson MJ, Curran MD, Court JA, Mann DMA, Ikeda S, Cookson MR, Hardy J, Allsop D: α-synuclein implicated in Parkinson's Disease is present in extracellular biological fluids, including human plasma. FASEB J 2003;17:1-16. https://doi.org/10.1096/fj.03-0098fje |
||||
| 21 Pooler AM, Phillips EC, Lau DH, Noble W, Hanger DP: Physiological release of endogenous tau is stimulated by neuronal activity. EMBO Rep 2013;14:389-394. https://doi.org/10.1038/embor.2013.15 |
||||
| 22 Ghaffari A, Li Y, Kilani RT, Ghahary A: 14-3-3 sigma associates with cell surface aminopeptidase N in the regulation of matrix metalloproteinase-1. J Cell Sci 2010; 123:2996-3005. https://doi.org/10.1242/jcs.069484 |
||||
| 23 Ostrerova N, Petrucelli L, Farrer M, Mehta N, Choi P, Hardy J, Wolozin B: α-synuclein Shares Physical and Functional Homology with 14-3-3 Proteins. J Neurosci 1999;19:5782-5791. https://doi.org/10.1523/JNEUROSCI.19-14-05782.1999 |
||||
| 24 Djelloul M, Holmqvist S, Boza-Serrano A, Azevedo C, Yeung MS, Goldwurm S, Frisen J, Deierborg T, Roybon L: Alpha-Synuclein Expression in the Oligodendrocyte Lineage: an In vitro and In vivo Study Using Rodent and Human Models. Stem Cell Rep 2015;5:174-184. https://doi.org/10.1016/j.stemcr.2015.07.002 |
||||
| 25 Andringa G, Du F, Chase NT, Bennet C: Mapping of Rat Brain Using the Synuclein-1 Monoclonal Antibody Reveals Somatodendritic Expression of α-Synuclein in Populations of Neurons Homologous to those Vulnerable to Lewy Body Formation in Human Synucleinopathies, J Neuropath Exp Neurol 2003; 62:1060-1075. https://doi.org/10.1093/jnen/62.10.1060 |
||||
| 26 Perez RG, Waymire JC, Lin E, Liu JJ, Guo F, Zigmond MJ: A Role for α-Synuclein in the Regulation of Dopamine Biosynthesis. J Neurosci 2002;22:3090-3099. https://doi.org/10.1523/JNEUROSCI.22-08-03090.2002 |
||||
| 27 Abeilovich A, Schitz Y, Fariñas I, Choi-Lundber D, Ho WH, Castillo PE, Shinksy N, Garcia Verdugo JM, Amanini M, Ryan A, Hynes M, Phillips H, Sulzer D, Rosenthal A: Mice Lacking a-Synuclein Display Functional Deficits in the Nigrostriatal Dopamine System. Neuron 2000;25:239-252. https://doi.org/10.1016/S0896-6273(00)80886-7 |
||||
| 28 Sotirou E, Vassilatis DK, Vila M, Stefanis L: Selective noradrenergic vulnerability in alpha-synuclein transgenic mice. Neurobiol Aging 2010;31:2103-2114. https://doi.org/10.1016/j.neurobiolaging.2008.11.010 |
||||
| 29 Kujacic M, Carlsson A: In vivo activity of tyrosine hydroxylase in rat adrenal glands following administration of quinpirole and dopamine. Eur J Phamacol 1995;278:9-15. https://doi.org/10.1016/0014-2999(95)00092-Y |
||||
| 30 Nagatsu T, Nakashima A, Ichinose H, Kobayashi K: Human tyrosine hydroxylase in Parkinson's disease and in related disorders. J Neural Transm 2019;126:397-409. https://doi.org/10.1007/s00702-018-1903-3 |
||||
| 31 Chandra S, Fornai F, Bae Kwon H, Yazdani U, Atasoy D, Liu X, Hammer RE, Battaglia G, German DC, Castillo PE, Sudhof TC: Double knockout mice for α- and β-synucleins: effect on synaptic functions. Proc Natl Acad Sci USA 2004;101:14966-1497. https://doi.org/10.1073/pnas.0406283101 |
||||
| 32 Bonaccorsi di Patti MC, Angiulli E, Casini A, Vaccaro R, Cioni C, Toni M: Synuclein Analysis in Adult Xenopus laevis. Int J Mol Sci 2022;23:6058. https://doi.org/10.3390/ijms23116058 |
||||
| 33 Khoshi A, Goodarzi G, Mohammadi R, Arezumand R, Moghbell M, Nayarlyan NB: Reducing effect of insulin resistance on alpha-synuclein gene expression in skeletal muscle. Diabetol Metab Syndr 2019;11:99. https://doi.org/10.1186/s13098-019-0499-6 |
||||
| 34 Wijesekara N, Ahrens R, Wu L, Langman T, Tandon A, Fraser PE: α-Synuclein Regulates Peripheral Insulin Secretion and Glucose Transport. Front Aging Neurosci 2021;13:665348. https://doi.org/10.3389/fnagi.2021.665348 |
||||
| 35 Rodriguez-Araujo G, Nakagami H, Hayashi H, Mori M, Shiuchi T, Minokoshi Y, Nakoaka Y, Takami Y, Komuro I, Morishita R, Kaneda Y: Alpha-synuclein elicits glucose uptake and utilization in adipocytes through the Gab1/PI3K/Akt transduction pathway. Cell Mol Life Sci 2013; 70:1123-1133. https://doi.org/10.1007/s00018-012-1198-8 |
||||
| 36 Vega-Moreno J, Tirado-Cortes A, Alvarez R, Irles C, Mas-Oliva J, Ortega A: Cholesterol depletion uncouples β-dystroglycans from discrete sarcolemmal domains, reducing the mechanical activity of skeletal muscle. Cell Physiol Biochem 2012;29:905-918. https://doi.org/10.1159/000186933 |
||||
| 37 Ortega A, Lepock RJ: Use of thermal analysis to distinguish magnesium and calcium stimulated ATPase activity in isolated transverse tubule membranes from skeletal muscle. Biochim Biophys Acta 1995;1233:7-13. https://doi.org/10.1016/0005-2736(94)00243-I |
||||
| 38 Jiménez-Garduño AM, Mitkovski M, Alexopoulos IK, Sanchez A, Stühmer W, Pardo LA, Ortega A: KV10.1 K (+)-channel plasma membrane discrete domain partitioning and its functional correlation in neurons. Biochim Biophys Acta 2014;1838:921-931. https://doi.org/10.1016/j.bbamem.2013.11.007 |
||||
| 39 Schindler J, Nothwang HG: Enrichment of brain plasma membranes by affinity two-phase partitioning. Methods Mol Biol 2009;528:119-126. https://doi.org/10.1007/978-1-60327-310-7_8 |
||||
| 40 Simons K, Ikonen E: Functional rafts in cell membranes. Nature 1997;387:569-572. https://doi.org/10.1038/42408 |
||||
| 41 Solares-Perez A, Alvarez R, Crosbie RH, Vega-Moreno J, Medina-Monares J, Estrada FJ, Ortega A, Coral-Vazquez R: Altered calcium pump and secondary deficiency of γ-sarcoglycan and microspan in sarcoplasmic reticulum membranes isolated from δ-sarcoglycan knockout mice. Cell Calcium 2010;48:28-36. https://doi.org/10.1016/j.ceca.2010.06.003 |
||||
| 42 Baltic S, Perovic M, Mladenovic A, Raicevic N, Ruzdijic S, Rakic L, Kanazir S: α-Synuclein is expressed in different tissues during human fetal development. J Mol Neurosci 2004;22:199-203. https://doi.org/10.1385/JMN:22:3:199 |
||||
| 43 Yu W, Yang W, Li X, Li X, Yu S: Alpha-synuclein oligomerization increases its effect on promoting NMDA receptor internalization. Int J Clin Exp Pathol 2019;12:87-100. | ||||
| 44 Delenclos M, Burgess JD, Lamprokostopoulou A, Outeiro TF, Vekrellis K, McLean JP: Cellular models of alpha-synuclein toxicity and aggregation. J Neurochem 2019;150:566-576. https://doi.org/10.1111/jnc.14806 |
||||
| 45 Bartels T, Choi J, Selkoe D: α-Synuclein occurs physiologically as a helically folded tetramer that resists aggregation. Nature 2011;477:107-110. https://doi.org/10.1038/nature10324 |
||||
| 46 Nakai M, Fujita M, Waragai M, Sugama S, Wei J, Akatsu H, Ohtaka-Mauyama C, Okado H, Hashimoto M: Expression of α-synuclein, a presynaptic protein implicated in Parkinson's disease, in erythropoietic lineage. Biochem Biophys Res Commun 2007;358:104-110. https://doi.org/10.1016/j.bbrc.2007.04.108 |
||||
| 47 Araki A, Sugawara K, Hayakawa EH, Ubukawa K, Kobayashi I, Wakui H, Takahashi N, Sawada K, Mochizuki H, Nunomura W: Delocalization of alpha Synuclein in the process or differentiation of human erythroid cells. Int J Hematol 2018;108:130-138. https://doi.org/10.1007/s12185-018-2457-8 |
||||
| 48 Wang W, Perovic I, Chittuluru J, Kaganovich A, Nguyen LTT, Liao J, Auclair JR, Johnson D, Landau A, Simorellis AK, Ju S, Cookson MR, Asturias FJ, Agar HJ, Web BM, Kang C, Ringe D, Petsko GA, Pochapsky TC, Hoang QQ: A soluble α-synuclein construct forms a dynamic tetramer. Proc Natl Acad Sci 2011;108:17797-17802. https://doi.org/10.1073/pnas.1113260108 |
||||
| 49 Tanaka M, Kim YM, Lee G, Junn E, Iwatsubo T, Mouradian MM: Aggresomes Formed by α-Synuclein and Synphilin-1 Are Cytoprotective. J Biol Chem 2003;279:4625-4631. https://doi.org/10.1074/jbc.M310994200 |
||||
| 50 Saudou F, Finkbeiner S, Devys D, Greenberg ME: Huntingtin acts in the nucleus to induce apoptosis but death does not correlate with the formation of intranuclear inclusions. Cell 1998; 95:55-66. https://doi.org/10.1016/S0092-8674(00)81782-1 |
||||
| 51 Arrasate M, Mitra S, Schweitzer ES, Segal MR, Finkbeiner S. Inclusion body formation reduces levels of mutant huntingtin and the risk of neuronal death. Nature 2004;431:85-810. https://doi.org/10.1038/nature02998 |
||||
| 52 Fujiwara H, Hasegawa M, Dohmae N, Kawashima A, Masliah E, Goldberg MS, Shen J, Takio K, Iwatsubo T: α-Synuclein is phosphorylated in synucleinopathy lesions. Nat Cell Biol 2002:4:160-164. https://doi.org/10.1038/ncb748 |
||||
| 53 Hu S, Hu M, Liu J, Zhang B, Zhang Z, Zhou FH, Wang L, Dong J. Phosphorylation of Tau and α-Synuclein Induced Neurodegeneration in MPTP Mouse Model of Parkinson's Disease. Neuropsychiatr Dis Treat 2020;16:651-663. https://doi.org/10.2147/NDT.S235562 |
||||
| 54 Wang Y, Shi M, Chung KA, Zabetian CP, Leverenz JB, Berg D, Srulijes K, Tojanowski JQ, Lee VM, Siderowf AD, Hurtig H, Litvan I, Schiess MC, Peskind ER, Masuda M, Hasegawa M, Lin X, Pan C, Galasko D, Goldstein DS, Jensen PH, Yang H, Cain KC, Zhang J: Phosphorylated α-Synuclein in Parkinson's Disease. Sci Transl Med 2012;4:121ra20. https://doi.org/10.1126/scitranslmed.3002566 |
||||
| 55 Fayyad M, Majbour NK, Vaikath NN, Erskine D, El-Tarawneh EH, Sudhakaran IP, Abdesselem H, El-Agnaf OMA. Generation of monoclonal antibodies against phosphorylated α-Synuclein at serine 129: Research tools for synucleinopathies. Neurosci Lett 2020;725:13489. https://doi.org/10.1016/j.neulet.2020.134899 |
||||
| 56 Sasaki A, Arawaka S, Sato H, Kato T: Sensitive western blotting for detection of endogenous Ser129-phosphorylated α-synuclein in intracellular and extracellular spaces. Sci Rep 2015;5:14211. https://doi.org/10.1038/srep14211 |
||||
| 57 Peachey LD: The sarcoplasmic reticulum and the transverse tubules of the frog's Sartorius. J Cell Biol 1965;25:209-231. https://doi.org/10.1083/jcb.25.3.209 |
||||
| 58 Williams KD, Smith OD: Cholesterol conservation in skeletal muscle associated with age- and denervation-related atrophy. Brain Res 1989;493:14-22. https://doi.org/10.1016/0006-8993(89)90995-5 |
||||
| 59 Crouse JR, Grundy SM, Ahrens EH Jr: Cholesterol distribution in the bulk tissues of man: variation with age. J Clin Invest 1972;51:1292-1296. https://doi.org/10.1172/JCI106924 |
||||
| 60 Zhang Y, Appelkvist E, Kristensson K, Dallner G: The Lipid Compositions of Different Regions of Rat Brain During Development and Aging. Neurobiol Aging 1996;17:869-875. https://doi.org/10.1016/S0197-4580(96)00076-0 |
||||
| 61 Fortin DL, Troyer MD, Nakamura K, Kubo S, Anthony MD, Edwards RH: Lipid rafts mediate the synaptic localization of alpha-synuclein. J Neurosci 2004;24:6715-6723. https://doi.org/10.1523/JNEUROSCI.1594-04.2004 |
||||
| 62 Lee HJ, Choi C, Lee SJ: Membrane bound α-synuclein has a high aggregation propensity and the ability to seed the aggregation of the cytosolic form. J Biol Chem 2002;277:671-678. https://doi.org/10.1074/jbc.M107045200 |
||||
| 63 Mulgrew-Nesbitt A, Diraviyam K, Wang J, Sings S, Murray P, Li Z, Rogers L, Mirkovic N, Murray D: The role of electrostatics in protein-membrane interactions. Biochim Biophys Acta 2006;1761:812-826. https://doi.org/10.1016/j.bbalip.2006.07.002 |
||||
| 64 Tsigelny IF, Sharikov Y, Wrasidlo W, Gonzalez T, Desplats PA, Crews L, Spencer B, Masliah E: Role of α-synuclein penetration into the membrane in the mechanisms of oligomer pore formation. FEBS J 2012; 279:1000-1013. https://doi.org/10.1111/j.1742-4658.2012.08489.x |
||||
| 65 Chandra S, Chen X, Rizo J, Jahn R, Südhof TC: A Broken α-Helix in Folded α-Synuclein. J Biol Chem 2002;278:15313-15318. https://doi.org/10.1074/jbc.M213128200 |
||||
| 66 Pitha J, Irie T, Sklar PB, Nye JS: Drug solubilizers to aid pharmacologists: amorphous cyclodextrin derivatives. Life Sci 1988;43:493-502, https://doi.org/10.1016/0024-3205(88)90150-6 |
||||
| 67 Ilangumaran S, Hoessli DC: Effects of cholesterol depletion by cyclodextrin on the sphingolipid microdomains of the plasma membrane. Biochem J 1998; 335:433-440. https://doi.org/10.1042/bj3350433 |
||||
| 68 Hanada K, Nishijima M, Akamatsu Y, Pagano RE: Both sphingolipids and cholesterol participate in the detergent insolubility of alkaline phosphatase, a glycosylphosphatidylinositol-anchored protein, in mammalian membranes. J Biol Chem 1995;270:6254-6260. https://doi.org/10.1074/jbc.270.11.6254 |
||||
| 69 Foster LJ, Hong CL, Mann M: Unbiased quantitative proteomics of lipid rafts reveals high specificity for signaling factors. Proc Natl Acad Sci U S A 2003;100:5813-5818. https://doi.org/10.1073/pnas.0631608100 |
||||
| 70 Stefanova N, Klimaschewski L, Poewe W, Wenning GK, Reindl M: Glial cell death induced by overexpression of alpha-synuclein. J Neurosci Res 2001;65:432-438. https://doi.org/10.1002/jnr.1171 |
||||
| 71 Erustes AG, Stefani FY, Terashima JY, Stilhano RS, Monteforte PT, Pereira G, Han SW, Calgarotto AK, Hsu Y, Ureshino RP, Bincoletto C: Overexpression of α-synuclein in an astrocyte cell line promotes autophagy inhibition and apoptosis. J Neuro Res 2017;96:160-171. https://doi.org/10.1002/jnr.24092 |
||||
| 72 Danzer KM, Krebs SK, Wolff M, Birk G, Hengerer B: Seeding induced by α-synuclein oligomers provides evidence for spreading of α-synuclein pathology. J Neurochem 2009;111:192-203. https://doi.org/10.1111/j.1471-4159.2009.06324.x |
||||
| 73 Ahn KJ, Paik SR, Chung KC, Kim J: Amino acid sequence motifs and mechanistic features of the membrane translocation of alpha-synuclein. J Neurochem 2006; 97:265-279. https://doi.org/10.1111/j.1471-4159.2006.03731.x |
||||
| 74 Sui Y, Bullock KM, Erickson MA, Zhang J, Banks WA: Alpha synuclein is transported into and out of the brain by the blood-brain barrier. Peptides 2014;62:197-202. https://doi.org/10.1016/j.peptides.2014.09.018 |
||||
| 75 Cefalu WT, Wang ZQ, Bell-Farrow AD, Terry JG, Sonntag W, Waite M, Parks J: Chronic caloric restriction alters muscle membrane fatty acid content. Exp Gerontol 2000;35:331-341. https://doi.org/10.1016/S0531-5565(00)00093-0 |
||||