×
![]()
Corresponding Author: Katarzyna Smolarz
Institute of Oceanography, Department of Marine Ecosystems Functioning
University of Gdańsk, Piłsudskiego 46, 81-378 Gdynia (Poland)
Tel. +48585236861, Fax +48585236878, E-Mail katarzyna.smolarz@ug.edu.pl
Bivalves Transmissible Neoplasia: Biochemical Aspects of Contagious Cancer in a Clam Macoma Balthica
Anna Hallmanna Alicja Michnowskab Agnieszka Chomiczewskaa Marcin Lipińskia Katarzyna Smolarzb
aFaculty of Pharmacy, Department of Pharmaceutical Biochemistry, Medical University of Gdańsk, Gdańsk, Poland, bInstitute of Oceanography, Department of Oceanography and Geography, University of Gdańsk, Gdynia, Poland
Introduction
Neoplasia is considered a prominent disease affecting various species of commercial and ecological importance [1–3]. Distinctive cell morphology (cellular and nuclear polymorphism, hypertrophy and anaplasia), high mitotic activity, and a high level of aneuploidy are typical for the disease [4, 5]. Bivalves disseminated neoplasia (DN), as it originates from unregulated cell division presumably of haemocyte precursors, is also called haemic neoplasia or leukaemia-like cancer [3]. Some characteristics of DN (immunosuppression, altered respiration pathways and energetic metabolism) were found to be similar to those typical for leukaemia cancers present in vertebrates. DN in marine bivalves have been documented since the 1960’s in around 30 species [1–3, 6]. Historically, a role of pollutants in genome destabilization and induction of the disease was suggested, but its aetiology remains unclear [7]. Most recent data suggest that DN may belong to diseases of infectious nature, possibly transmissible by cohabitation of healthy and neoplastic animals in large bivalve assemblages [8, 9]. Indeed, various molecular analysis revealed that some DN’s have been identified as transmissible cancers in several bivalve species thus this type of a disease is called bivalves transmissible neoplasia (BTN). Although ten BTN lineages have been described to date [10–16], much investigation is still needed to know how many and which DN have a transmissible origin.
The disease was also found to occur in the Baltic clam Macoma balthica (previously known as Limecola balthica), and was diagnosed in various clam populations inhabiting different parts of the Northern Europe, e.g.: Wadden Sea [17], Finnish Baltic coast [18] and the most comprehensive set of research concern Southern Baltic populations from Polish coastal zone [5, 19, 20]. The possible transmission of independent cancer lineages was recently studied as a factor responsible for the high frequency of neoplasia in M. balthica from the Gulf of Gdańsk [16]. In cases of BTN, genotypes of neoplastic cells are nearly identical and they do not match those of the solid tissue of the host animal with different alleles dominating in normal and neoplastic cells. Indeed, published data on mitochondrial cytochrome c oxidase I (mtCOI ) sequences and nuclear elongation factor 1α (EF1α ) highlighted unique and divergent genotype of neoplastic cells and confirmed high similarity between DN cases obtained from different M. balthica clams. Thus, the contagious nature of the disease and the occurrence of BTN in M. balthica inhabiting the Gulf of Gdańsk were confirmed [16].
Do to the relatively often occurrence of the disease in the southern Baltic Sea population of the species, M. balthica appears as a perfect model for studying various, still unknown, physiological and biochemical aspects of BTN. Researching the disease using clams population suffering from BTN in natural conditions opens a new avenue not only for studying cancer epidemiology in wildlife, but also for using BTN as a model in cancer research. That includes cancer pathogenicity, energy metabolism or the ability to affect the new host.
In neoplastic disorders, the Warburg effect is a commonly accepted theory explaining cancer energy metabolism. The Warburg effect shifts the metabolic pathway of ATP generation in cancer cells from mitochondrial oxidative phosphorylation to anaerobic glycolysis and production of lactate. Hence, mitochondrial energetic functions of those cells are suppressed, and, as a consequence, decreasing activities of electron transport complexes and lowered oxygen consumption by mitochondria occur [21]. Indeed, according to Galuzzi et al. [22] formation of ATP in cancer cells is coupled to comparatively low levels of oxidative phosphorylation. Recent findings highlighted that in some leukemic disorders leukemic cells lose their capacity for mitochondrial respiration at the level of complex I (NADH-coenzyme Q reductase) and complex II (succinate-coenzyme Q reductase), the major sites for entry of electrons into the respiratory chain [23]. However, despite being an extremely important factor potentially boosting cancer pathogenicity, energy metabolism in DN, and in particular BTN, is unknown. Other important end products, substrates and/or intermediates of anaerobic energy metabolism supporting crucial life functions are free amino acids (FAA) [24]. It is a common knowledge that FAA levels in the tissues of aquatic organisms change not only seasonally, but also as a response to environmental stress or worsening physiological conditions [25, 26]. Yet, there is no data about FAA levels in haemolymph of bivalves suffering from either BTN or DN. Due to high mitotic and metabolic activities of DN/BTN cells, the working hypothesis here was that changes in FAA content, in particular in glutamine (Gln) level, point to the presence of additional source of energy for use and storage at the cellular level able helping to satisfy their elevated energy requirements.
Steroids are naturally made by a variety of living organisms as they help controlling many functions including the immune system, reproduction, inflammatory response and/or metabolism. Corticosteroids are steroid hormones that regulate metabolism of proteins, carbohydrates and fats, which is of crucial importance for the body’s energy homeostasis. They are also known for their anti-inflammatory and immunosuppressive abilities [27]. The final metabolites of the corticosteroid synthesis pathway include dehydrocorticosterone (D-CRT), corticosterone (CRT), cortisone (CN) and cortisol (HCN). However, synthesis and hormonal activity of HCN has so far been documented mainly in Primates, while CRT is known to occur in other animal species. In marine bivalves, the levels, the role and the origin of corticosteroids (glucocorticoids in particular) are not well understood, particularly in neoplastic disorders. Also, altered steroids levels may indicate an important role of this hormones in cancer biochemistry and their possible role in immunosuppression of the host.
As the occurrence of BTN in M. balthica population studied here was confirmed [16], we present the first data describing the physiology and biochemistry of BTN contrasted with those measured in healthy clams. Selected markers included glycolytic capacity, ROS production, FAA levels and enzymatic activity. As the BTN cells, after entering the host, must bypass its immune system, we studied if there are changes in signalling molecules such as corticosteroids (also as they have the ability to reduce inflammatory processes) in DN positive and negative clams. As BTN cells are characterised by elevated mitotic and metabolic activities and a high potential to metastasise, we looked at cellular respiration processes to highlight potential changes in DN cells able to satisfy their elevated energy requirements. Thus, the purpose of our work was to analyse physiological and biochemical aspects of DN/BTN in the Baltic clam M. balthica that can increase our knowledge about environmental carcinogenesis.
Materials and Methods
M. balthica was collected onboard of R/V Oceanograf using a benthic sledge in the Gulf of Gdańsk (Baltic Sea, Poland) at a sampling location (18°50.5’E 54°36.2’N) where BTN has recently been confirmed to occur in the clams’ inhabiting the area [16]. Thus, despite the fact that in this work we used physiological diagnostic tools to distinguish between DN and healthy clams, DN occurring in studied here clam population was proven to have a transmissible origin, and will be further called DN-BTN or BTN. After transportation to the laboratory, clams were opened, and haemolymph samples were collected from the pericardium or adductor muscle using microliter glass syringes. Preliminary neoplasia diagnosis was performed under light microscope based on haemolymph subsample stained with 0,5% methylene blue. For further analyses 46 healthy and all (in total 40) with advanced DN-BTN clams were selected. The latter ones were characterised by strong domination of neoplastic over normal haemocytes in the haemolymph sample with over 90% of abnormal haemocytes present in the haemolymph sample. Individuals at early DN-BTN stages were not taken into consideration in this study. Selected clams were treated according to three main protocols as described below. The first protocol included measurement of FAA levels (section 2.1) in the haemolymph. After collecting haemolymph sample, remaining soft tissue was stored in Davidson fixative for 48 h, transferred to 4% buffered formaldehyde solution, dehydrated, impregnated in xylene, embedded in paraffin and sectioned using a semi-automatic microtome on 3µm thick sections. Such prepared semi-thin sections were re-hydrated and stained with H&E using basic histology protocols. This set of samples (12 healthy and 11 neoplastic individuals) was used for final diagnosis of DN-BTN and for assessing the accuracy of preliminary diagnosis based on haemolymph subsample (Fig. 1A-E). Second protocol included measurement of mitochondrial respiration in 18 healthy and 13 neoplastic individuals. Here, mitochondria were isolated from the haemolymph sample while the soft tissue was homogenized and further processed as described in the sections 2.2.1 and 2.2.2 Third protocol (section 2.3.) included an assessment of the antioxidative response and measurement of biomarkers of physiological stress. For that purpose, after diagnosis, the remaining soft tissue was immediately frozen in liquid nitrogen and further stored in -80 °C until described in the section 2.1 analyses were performed. Here, 6 healthy and 6 neoplastic clams were selected. Final protocol was designed for analyses of corticosteroids levels performed based on 10 healthy and 10 neoplastic clams (section 2.4). A table showing the numbers of individuals used for each analysis is presented in the Supplementary Material (Supplementary Table S1 – for all supplementary material see www.cellphysiolbiochem.com).
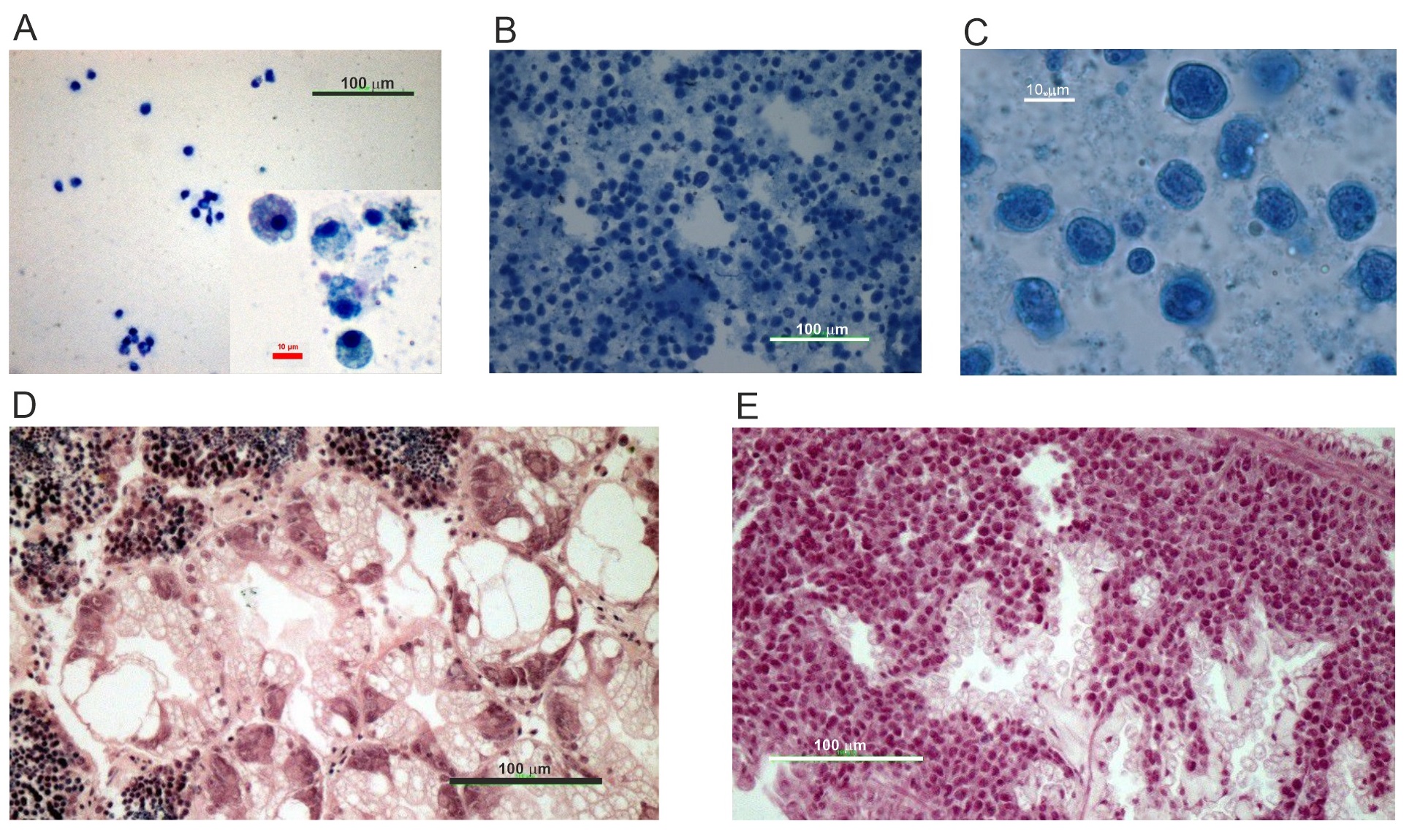
Free Amino Acids (FAA) in haemolymph samples
FAA concentrations were determined according to Olkowicz et al. [28]. 10 µL of the internal standard solution (2-chloroadenosine) was added to 50 µL of haemolymph (without haemocytes), and the mix was extracted with 120 µL of acetonitrile. Samples were precipitated for 30 minutes in ice, centrifuged 14 000 x g at 4◦C for 20 min., the supernatant was transferred into new tubes and freeze dried. The samples were further dissolved in 100 µL water, centrifuged for 15 min at 14 000 x g, 4°C and analysed using high performance liquid chromatography-mass spectrometry (LC/MS) on a Surveyor HPLC system coupled with a TSQ Vantage Triple-Stage Quadrupole mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA). Heated electrospray ionization in positive mode was used. Chromatographic separation was achieved with a 50 x 2 mm Synergi Hydro-RP 100 column with a 2.5 µm particle size (Phenomenex, Torrance, CA, USA). The mobile phase consisted of water with 5 mM nonafluoropentanoic acid (Buffer A) and acetonitrile with 0.1% formic acid (Buffer B). 2 µL aliquots of samples were injected into a column eluted with a mobile phase at a flow rate of 0.2 mL/min. Based on the retention times of individual amino acid standards, the identities of amino acids in the samples were determined and compared to the peaks of corresponding standards. The procedure was followed by calculation of quantities of individual amino acids.
Measurement of mitochondrial respiration and enzymatic activities
Used reagents include substrates: succinic acid, malic acid, pyruvic acid, ADP, inhibitors: rotenone (complex I inhibitor), antimycin A (complex III inhibitor), oligomycin (ATPase inhibitor), and ascorbate with N1,N1,N1,N1-tetramethyl-1,4-phenylene diamine (TMPD) as electron acceptor, carbonylcyanide-3-chlorophenylhydra-zone (CCCP) as ionophore (uncoupler).
Mitochondrial isolation. Mitochondria were isolated according to Schnaitman and Greenawalt [29] with some modifications. Single clam soft tissue was homogenised in 1 ml of MSHE buffer (210 mM mannitol, 70 mM sucrose, 5 mM HEPES, 1 mM EGTA and 0.5% BSA (fatty acid free), pH 7.2) at 4°C using glass homogenizer. Homogenate was centrifuged at 800 x g for 10 minutes at 4°C, obtained supernatant was centrifuged at 8000 x g for 15 minutes at 4°C and the pellet was resuspended in MSHE buffer. Total mitochondrial protein (mg/mL) was determined using the Bradford Assay reagent (Bio-Rad) and Bradford [30] method.
Mitochondrial respiration . Mitochondrial respiration was measured by the Agilent Seahorse XFp analyzer (Agilent Technologies, MA, USA). For analysis of mitochondrial coupling efficiency, the plate containing 25 µg mitochondria per well in 25 µL mitochondrial assay buffer (MAS; 220 mM mannitol, 70 mM sucrose, 10 mM KH2PO4, 5 mM MgCl2, 2 mM HEPES, 1mM EGTA and 0.2% BSA (fatty acid free), pH 7.2) was centrifuged (2000 x g, 15 min, 4°C) and 155 µl MAS containing 10 mM succinate pre-warmed to 20◦C was used. After calibration, the mitochondria plate was inserted. After 10 min, two cycles (1 min mixing, 3 min waiting time) were followed by three cycles (1 min mixing, 1 min waiting, 3 min measurement). The protocol applied before each injection included 2 min mixing and 1 min waiting, addition of the indicated compound, 1 min waiting and a final measurement (3 min). At the indicated time points, 4 mM ADP, 2.5 µg/mL oligomycin A, 4 µM CCCP and 4 µM antimycin A were added.
For analysis of the electron transport chain (ETC) complexes activities the plate containing 25 µg mitochondria per well in 25 µL MAS buffer was centrifuged (2000 x g, 15 min, 4°C) and 155 µl MAS buffer containing 10 mM pyruvate, 2 mM malate and 4 µM CCCP prewarmed to 20◦C were used. In the same time sequence as in mitochondrial coupling assay, at the indicated time points, final concentrations of 2 µM rotenone, 10 mM succinate, 4 µM antimycin A and 10mM ascorbate plus 100 µM TMPD were added. Oxygen consumption rates (OCR; pmol/min) were calculated by the Wave software (Seahorse Bioscience).
Mitochondrial enzymes activities assay. The succinate dehydrogenase (SDH, EC 1.3.5.1) activity was determined in isolated bivalve mitochondria using a commercially available test kit (MAK197, Sigma-Aldrich). SDH catalyzes the oxidation of succinate to fumarate and carries electrons from FADH to CoQ. SDH activity was determined by generating a product (commercial DCIP) of which absorbance at 600 nm was proportional to the observed enzymatic activity. SDH activity was expressed in nmol/min/mg of the mitochondrial protein.
The cytochrome c oxidase (COX, EC 1.9.3.1) activity was determined using a commercially available test kit (CYTOCOX1, Sigma-Aldrich). COX is the principal terminal oxidase of high affinity oxygen in the aerobic metabolism of all animals. The activity of COX was measured based on its decrease in absorbance at 550 nm of ferrocytochrome c caused by its oxidation to ferricytochrome c by cytochrome c oxidase. COX activity was expressed in nmol/min/mg of the mitochondrial protein.
Antioxidative and physiological stress biomarkers
Individual organisms were homogenized manually with a Teflon-pestle homogenizer in ice-cold buffer (to obtain 20% homogenate) containing 50 mM Tris-H2SO4, pH 7.6 with 0.1 mM EDTA, 1 mM PMSF, 2 mM DTT and 0.2% Triton X-100. The homogenates were centrifuged at 14 000 x g for 30 minutes at 4°C using Centrifuge SIGMA 3K18. The supernatants were transferred into fresh tubes and used for analysis. Cytosolic protein was determined by Lowry method [31] with modification of Peterson [32].
The TAC (Total Antioxidant Capacity) assay was measured using Total Antioxidant Capacity Assay Kit (MAK187, Sigma-Aldrich). Concentrations of small molecule and protein antioxidants (e.g. tocopherols, carotenes, vitamin A, ubiquinols) were determined using Trolox, a water-soluble vitamin E analog serving as an antioxidant standard. The prepared samples were measured at 570 nm in Synergy 2 Multi-Mode Reader (BioTek). The amount of TAC was normalized to the protein content of the samples and expressed in nmol/mg of total protein concentration.
Acetylcholinesterase (ACHE; EC 3.1.1.7) activity was measured with AChE Activity Assay Kit (MAK119, Sigma-Aldrich). This assay is an optimized version of the Ellman method in which thiocholine, produced by AChE, reacts with 5,5’-dithiobis(2-nitrobenzoic acid) to form an colorimetric product, proportional to the AChE activity present. The absorbance was measured for 10 min at 412 nm in Synergy 2 Multi-Mode Reader (BioTek). AChE activity was expressed in nmol/min/mg of total protein concentration.
Glutathione S-transferase (GST; EC 2.5.1.18) activity was determined spectrophotometrically (UV-VIS Spectrophotometer, Beckman Coulter) by the method of Habig et al. [33]. The reaction mixture contained 100 mM phosphate-buffered saline buffer (pH 6.5), 100 mM 1-chloro-2,4-dinitrobenzene (CDNB) and the reaction started by adding supernatant and 100 mM glutathione as substrate. The absorbance was measured for 5 min at 340 nm. GST activity was expressed in nmol/min/mg of total protein concentration.
CBO level was detected using the Protein Carbonyl Content Assay Kit (MAK094, Sigma-Aldrich). Carbonyl content was determined by the derivatization of protein carbonyl groups with 2,4-dinitrophenylhydrazine (DNPH) leading to the formation of stable dinitrophenyl (DNP) hydrazone adducts that are proportional to the carbonyls present. Absorbance was measured at 375 nm in Synergy 2 Multi-Mode Reader (BioTek). The amount of CBO was normalized to the protein content of the samples and expressed in nmol/mg of total protein concentration.
Malondialdehyde (MDA) level was detected using the Lipid Peroxidation (MDA) Assay Kit (MAK085, Sigma-Aldrich) by the reaction of MDA with thiobarbituric acid (TBA) forming amount of colorimetric product proportional to MDA. Samples were pipetted on a 96 - well microplate and absorbance was measured at 532 nm in Synergy 2 Multi-Mode Reader (BioTek). The amount of MDA was normalized to the samples protein content and expressed in nmol/mg of total protein concentration.
Determination of corticosteroids in M. balthica tissues
Corticosteroids concentration was measured according to Jakubowska et al. [34]. Each clam was homogenized using 5 ml of buffer containing 8.5 mM MgCl2, 3.13 mM KCl, 7.59 mM NaCl, 2.7 mM CaCl2, 50 mM Tris/HCl, pH 7.4) per tissue gram. Next, MTBE (tert-butyl methyl ether) was added, the sample was mixed for 1 min and the mixture was centrifuged at 3000 × g for 15 min at room temperature. The supernatant was transferred to a new glass vial and MTBE was allowed to evaporate to dryness at 45°C. Upon evaporation, the dry residue of the eluate was dissolved in 100 μl of methanol and 5 μl of this solution was injected into the HPLC–ESI-MS/MS system. The elution was performed under a 10-min gradient. Testosterone-2,3,4-13C3 and 4-Androstene-3,17-dione-2, 3,4-13C3 were used as internal standards. HPLC-ESI-MS/MS analysis was performed using a triple-quadrupole mass spectrometer (TSQ Vantage, Thermo Scientific) equipped with an ion electrospray source (ESI) coupled with a HPLC system (Thermo Scientific) using selected reaction monitoring (SRM) for detection in positive ion mode. All analytes were detected in SRM. The columns Kinetex Phenyl-Hexyl 50 × 2.1 mm 1.7u and Kinetex Biphenyl 50 × 1 mm 1.7u (Phenomenex) were used. Buffers used for the HPLC gradient elution included buffor A (0.1% formic acid in deionized water) and B (0.1% formic acid in methanol). Column equilibration was performed for 10 min. The total analysis time was 10 min. Calibration curves were prepared based on matrix and plotted as the peak area ratio (standard/IS) versus the amount of analytes. Next, the quantity of selected corticosteroids was calculated.
Statistical analysis
Data were tested for normality with Shapiro–Wilk test. Statistical significance of differences between groups (healthy and DN-BTN) was verified with Kruskal-Wallis test (non-parametric ANOVA) for data with non-normal distribution. The differences were considered significant at p<0.05. The results are presented as means ± standard errors of the mean (S.E.M). Calculations were performed using STATISTICA 13 software, figures were prepared in Sigma-Plot 11 software.
Results
FAA concentration
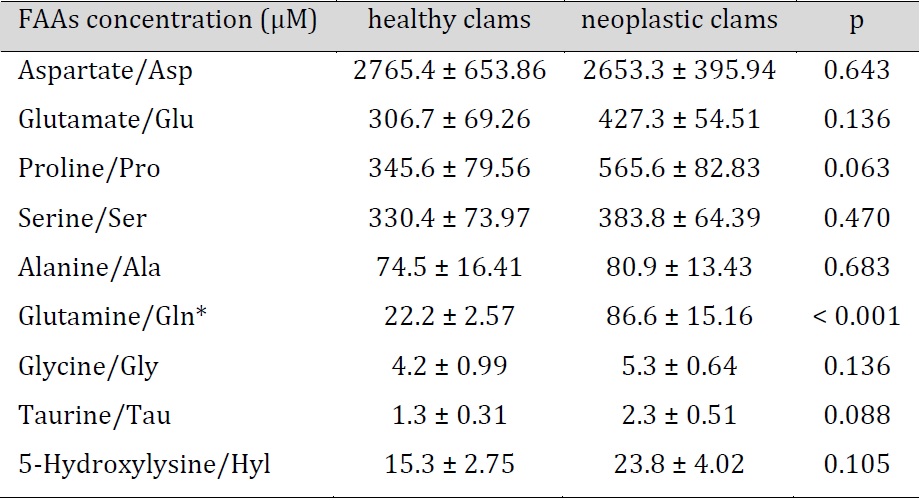
In general, nine amino acids were determined in bivalve hae-molymph (Table 1). Aspartate (Asp) was present at the highest concentration (about 2.7 mM) and taurine (Tau) was found at the lowest level (about 1.3 µM). Overall, the comparative analysis showed no differences between FAA levels in haemolymph of healthy and neoplastic clams, except for glutamine (Gln), which occurred at a significantly higher concentration in neoplastic (86.6 µM) when compared to healthy clams (22.2 µM).
Mitochondrial respiration and mitochondrial enzyme activities
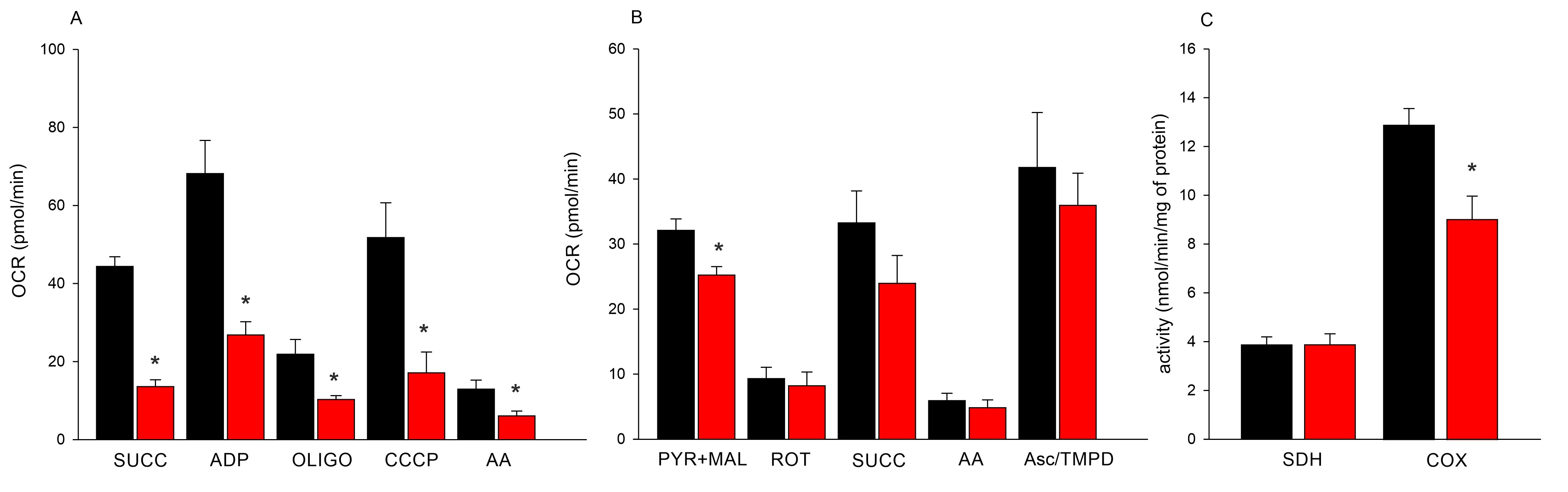
Coupled mitochondria iso-lated from neoplastic bivalves were characterised by decreased respiration efficiency when com-pared to those isolated from healthy clams. Mitochondrial oxygen consumption in the presence of succinate, as well as reduced respiratory induction in the presence of ADP, were 3 and 2.5 times lower in neoplastic than in healthy clams, respectively. Additionally, mitochondria iso-lated from DN-BTN bivalves were found to be less sensitive to used inhibitors in comparison to healthy ones (Fig. 2A). Examination of individual components of the respiratory chain in uncoupled mitochondria showed similar activity of respiratory chain proteins in the mitochondria of healthy and DN-BTN bivalves, except for complex I, in which the oxygen consumption capacity was significantly higher in healthy clams (Fig. 2B). Sensitivity of respiratory chain complexes to inhibitors such as rotenone, antimycin A and oligomycin was observed in the mitochondria of healthy and neoplastic bivalves. Moreover, the transfer of electrons through ETC to the artificial acceptor ascorbate/TMPD was observed in both clam groups. Mitochondrial SDH activity was low (3.9 nmol/min/mg of protein) and comparable in both groups (Fig. 2C). Mitochondrial cytochrome c oxidase activity was significantly lower in BTN-positive individuals (9.1 ± 1.0) in comparison to the healthy ones (12.9 ± 0.7 nmol/min/mg) (Fig. 2C).
Antioxidative and physiological stress biomarkers
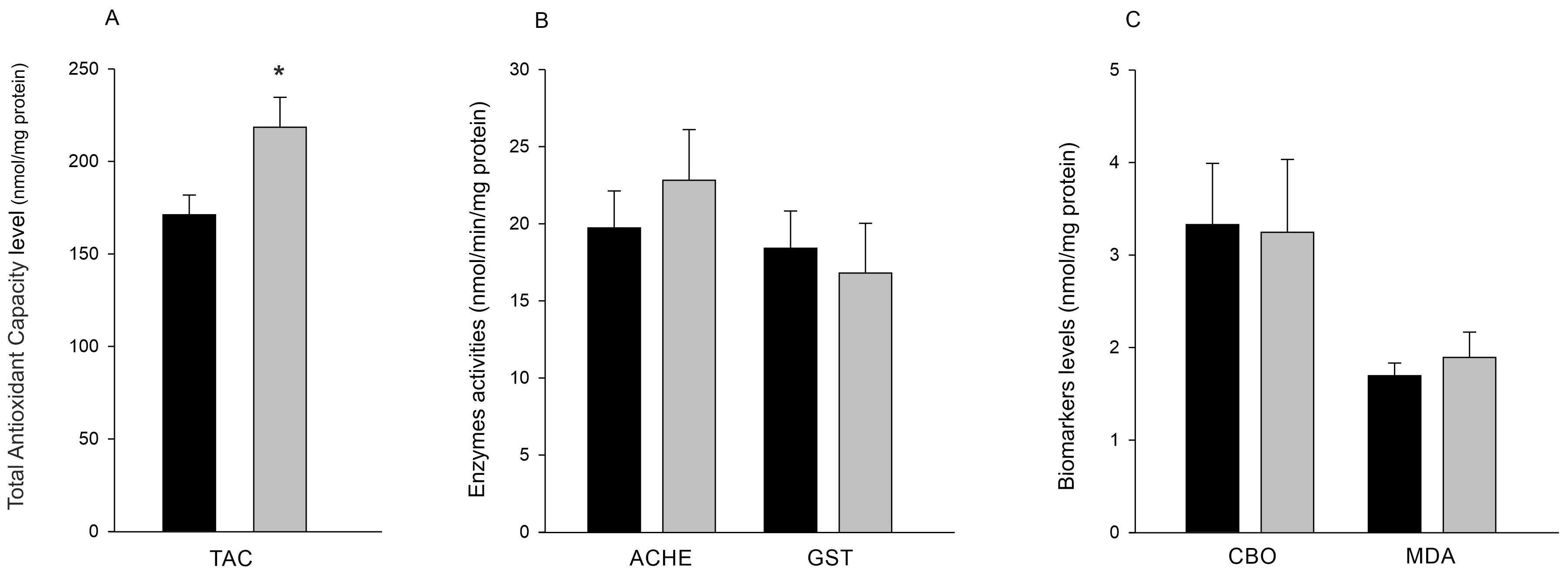
BTN-positive clams were characterised by significantly higher low-molecular antioxidant concentration then healthy ones (218.9 ±36.1 nmol/mg vs 171.3 ± 23.7 nmol/mg, p=0.028, Fig. 3A). AChE activity in tissues from healthy and BTN clams was at a similar level, 19.8 ± 2.4 and 22.8 ± 3.3 nmol/min/mg protein, respectively (Fig. 3B). Also, no statistically significant differences between healthy and BTN clams in GST activity were observed (18.5 ± 2.4 nmol/min/mg protein in healthy and 16.9 ± 3.2 nmol/min/mg protein in BTN clams, Fig. 3B). No statistically significant differences in the level of accumulation of protein peroxidation (CBO) and lipid peroxidation (MDA) products between healthy and neoplastic bivalves were found. The amount of CBO in healthy clams was 3.3 ± 0.7 nmol/mg protein while in BTN clams 3.2 ± 0.8 nmol/mg protein. The amount of MDA in healthy clams was 1.7 ± 0.1 nmol/mg protein while in those affected by BTN 1.9 ± 0.3 nmol/mg protein (Fig. 3C).
Corticosteroids
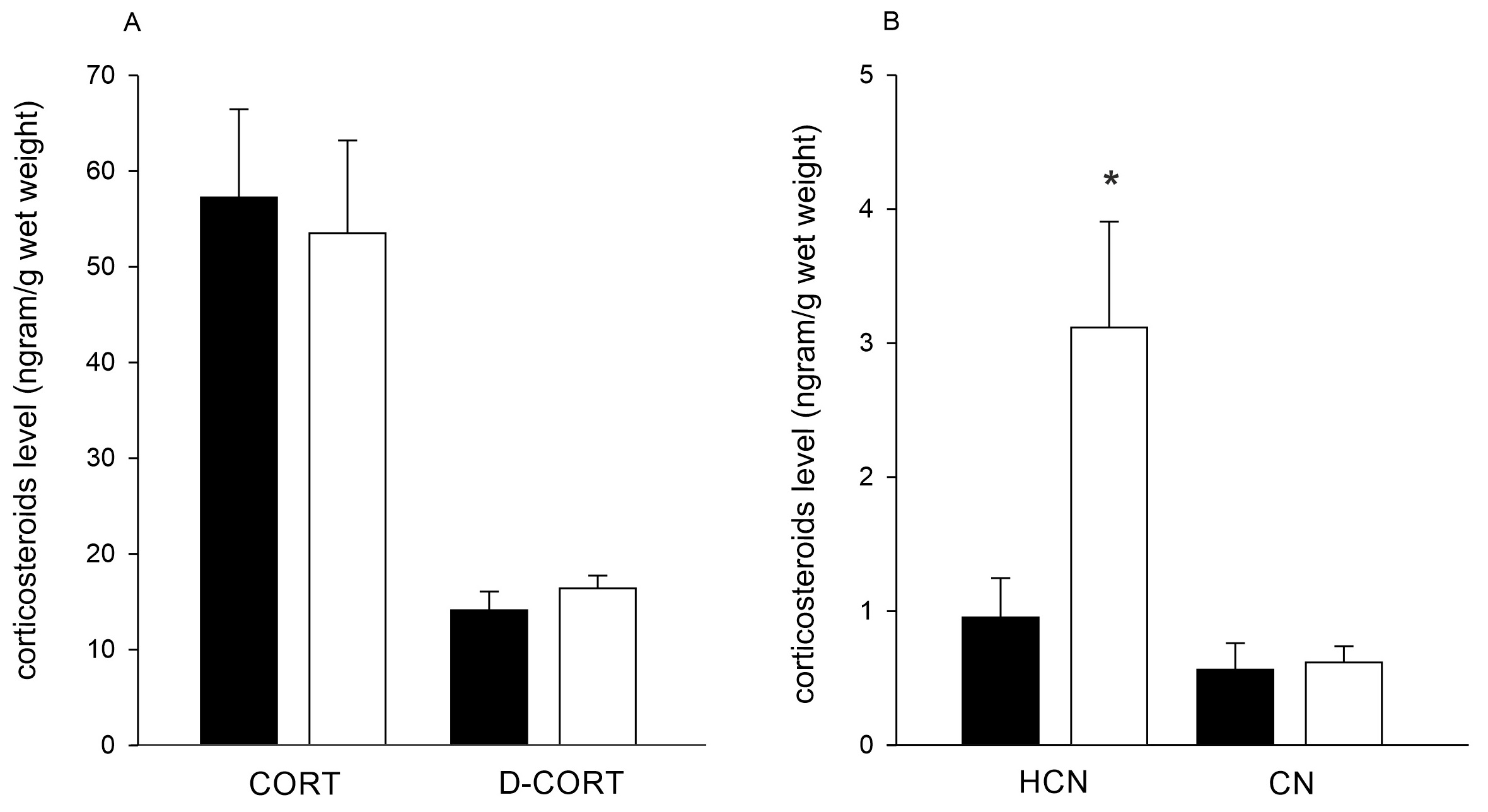
Corticosterone was found at similar concentration in both healthy (57.2 ± 9.2 ng/g w.w.) and DN-BTN (53.4 ± 9.7 ng/g w.w.) clams. Levels of dehydrocorticosterone were lower and comparable between both clam groups (14.1 ± 2.0 ng/g w.w. and 16.4 ± 1.3 ng/g w.w. in healthy and BTN clams, respectively, Fig. 4A). Cortisol was found at a concentration of 0.95 ± 0.3 ng/gram w.w. in healthy clams, and its level was significantly higher in BTN-positive clams (3.1 ± 0.8 ng/g w.w., p=0.005) (Fig. 4B). Cortisone was at the lowest level of all corticosteroids measured, and its concentration was similar in both clam groups (0.6 ± 0.2 ng/gram w.w. in healthy and 0.6 ± 0.1 ng/gram w.w. in BTN clams) (Fig. 4B).
Conclusion
Disseminated neoplasia, and in particular bivalve transmissible neoplasia, is a disease of unknown biochemical characteristics occurring in various bivalve species. To increase our knowledge about this phenomenon, we described FAA levels in bivalve haemolymph, corticosteroids levels, oxidative and physiological stress biomarkers response in clam tissue and measured mitochondrial respiration in DN-positive and healthy cells. In both clam groups nine FAAs were found (Asp, Glu, Pro, Ser, Ala, Gln, Gly, Tau and 5-hydroxylysine) but in the neoplastic clams significantly higher content of Gln was observed when compared to the healthy ones. Such high levels of Gln suggest its essential role in energy production in leukaemia-like cancer and stresses a potential role of Gln as a marker of environmental carcinogenesis. Analyses of the respiratory pathway highlighted significantly lower oxygen consumption in coupled and uncoupled mitochondria (respiratory complex I) isolated from DN-positive clams that indicated altered mitochondrial function in BTN. As an occurring Warburg effect reduces effectiveness of mitochondrial respiration, a switch to anaerobic metabolism and glutaminolysis was expected. Indeed, significantly lower cytochrome c oxidase activity was found in the DN-BTN-positive clams, potentially confirming their impaired mitochondrial respiration, and detectable activity of succinate dehydrogenase suggested the occurrence of aerobic metabolism. Indeed, an increased level of Gln highlights glutaminolysis as a potential energy source for BTN cells. Also, a significantly higher concentration of low molecular weight antioxidants level was confirmed in DN-BTN clams when compared to the healthy ones, but no elevated oxidative stress markers (MDA and CBO levels and GST activity) were found, suggesting an efficient antioxidant response. Finally, high levels of corticosterone and lower amounts of dehydrocorticosterone, cortisol and cortisone were present in healthy bivalves, while in the DN-BTN ones significantly elevated level of cortisol was found. As bivalve haemocytes belong to the main neuro-endocrine secretion systems, these changes may be explained by haemocytic activation and/or massive changes occurring in BTN.
Finalising, based on our results we are able to select three biomarkers as potential candidates for standardized biochemical markers for DB-BTN, namely Gln level, OCR rate and cortisol concentration. Yet, our study is the first one describing various biochemical features of the disease, thus more studies are needed to provide more comparative data regarding this aspect of the disease. That is in particular important as some of the measured features, such as FAA level, are known to change in a response not only to the physiological state of an individual, but also in response to the external environmental conditions.
We would also like to thank Dr. Michael Metzger from the Pacific Northwest Research Institute (Seattle, U.S.A.) for reviewing and commenting on the manuscript and language correction and Alicja Braczko from Medical University of Gdansk for help with mitochondrial analysis using Agilent Seahorse XFp analyser.
Author Contributions
Anna Hallmann – conceptualisation, analysis, data acquiring and interpretation, writing the original draft.
Alicja Michnowska – field sampling, analysis, data acquiring and interpretation, manuscript reviewing and editing.
Agnieszka Chomiczewska – analysis and data acquiring.
Marcin Lipiński – analysis and data acquiring.
Katarzyna Smolarz – funds provider, field sampling, analysis, data interpretation, manuscript writing, reviewing and editing.
Funding Sources
The analyses performed within this study were financed by the Harmonia grant 2017/26/M/NZ8/00478 from the Polish Ministry of Science and Higher Education.
Statement of Ethics
The authors confirm that the study was conducted ethically. There are no regulations considering using lower invertebrates, such as clams, for scientific purposes. However, the authors made sure to provide as little harm as possible to the studied animals.
The authors have no conflicts of interest to declare.
| 1 Díaz S, Renault T, Villalba A, Carballal M: Disseminated neoplasia in cockles Cerastoderma edule : ultrastructural characterisation and effects on haemolymph cell parameters. Dis Aquat Organ 2011;96:157-167. https://doi.org/10.3354/dao02384 |
||||
| 2 Gombač M, Sitar R, Pogačnik M, Arzul I, Jenčič V: Haemocytic neoplasia in Mediterranean mussels (Mytilus galloprovincialis ) in the Slovene Adriatic Sea. Mar Freshw Behav Physiol 2013;46:135-143. https://doi.org/10.1080/10236244.2013.782736 |
||||
| 3 Barber BJ: Neoplastic diseases of commercially important marine bivalves. Aquat Living Resour 2004;17:449-466. https://doi.org/10.1051/alr:2004052 |
||||
| 4 Smolarz K, Renault T, Soletchnik P, Wolowicz M: Neoplasia detection in Macoma balthica from the Gulf of Gdansk: Comparison of flow cytometry, histology and chromosome analysis. Dis Aquat Organ 2005;65:187-195. https://doi.org/10.3354/dao065187 |
||||
| 5 Smolarz K, Renault T, Soletchnik P, Wolowicz M: Survey for neoplasia in Macoma balthica from the Gulf of Gdansk by flow cytometry. Dis Aquat Organ 2005;66:41-46. https://doi.org/10.3354/dao066041 |
||||
| 6 Wołowicz M, Smolarz K, Sokołowski A: Neoplasia in Estuarine Bivalves: Effect of Feeding Behaviour and Pollution in the Gulf of Gdansk (Baltic Sea, Poland); in: Dame RF, Olenin S (eds): The Comparative Roles of Suspension-Feeders in Ecosystems. NATO Science Series IV: Earth and Environmental Series, vol 47. Springer, Dordrecht, 2005, pp 165-182. https://doi.org/10.1007/1-4020-3030-4_10 |
||||
| 7 Pariseau J, Saint-Louis R, Delaporte M, el Khair MA, McKenna P, Tremblay R, DavidsonT, Pelletier E, Berthe F: Potential link between exposure to fungicides chlorothalonil and mancozeb and haemic neoplasia development in the soft-shell clam Mya arenaria : A laboratory experiment. Mar Pollut Bull 2009;58:503-514. https://doi.org/10.1016/j.marpolbul.2008.12.011 |
||||
| 8 Mateo DR, MacCallum GS, Davidson J: Field and laboratory transmission studies of haemic neoplasia in the soft-shell clam, Mya arenaria , from Atlantic Canada. J Fish Dis 2016;39:913-927. https://doi.org/10.1111/jfd.12426 |
||||
| 9 Elston RA, Kent ML, Drum AS: Progression, lethality and remission of hemic neoplasia in the bay mussel Mytilus edulis . Dis Aquat Org 1988;4:135-142 https://doi.org/10.3354/dao004135 |
||||
| 10 Metzger MJ, Reinisch C, Sherry J, Goff SP: Horizontal transmission of clonal cancer cells causes leukemia in soft-shell clams. Cell 2015; 161:255-263. https://doi.org/10.1016/j.cell.2015.02.042 |
||||
| 11 Metzger MJ, Villalba A, Carballal MJ, Iglesias D, Sherry J, Reinisch C, Muttray A, Baldwin S, Goff S: Widespread transmission of independent cancer lineages within multiple bivalve species. Nature 2016;534:705-709. https://doi.org/10.1038/nature18599 |
||||
| 12 Garcia-Souto D, Bruzos AL, Diaz S, Rocha S, Pequeño-Valtierra A, Roman-Lewis CF, Alonso J, Rodriguez R, Costas D, Rodriguez-Castro J, Villanueva A, Silva L, Valencia JM, Annona G, Tarallo A, Ricardo F, Bratoš Cetinić A, Posada D, Pasantes JJ, Tubio JMC: Mitochondrial genome sequencing of marine leukaemias reveals cancer contagion between clam species in the Seas of Southern Europe. Elife 2022;11:e66946. https://doi.org/10.7554/eLife.66946 |
||||
| 13 Yonemitsu MA, Giersch RM, Polo-Prieto M, Hammel M, Simon A, Cremonte F, Avilés FT, Merino-Véliz N, Burioli EAV, Muttray AF, Sherry J, Reinisch C, Baldwin SA, Goff SP, Houssin M, Arriagada G, Vázquez N, Bierne N, Metzger MJ: A single clonal lineage of transmissible cancer identified in two marine mussel species in South America and Europe. Elife. 2019;8:47788. https://doi.org/10.7554/eLife.47788 |
||||
| 14 Skazina M, Odintsova N, Maiorova M, Ivanova A, Väinölä R, Strelkov P: First description of a widespread Mytilus trossulus -derived bivalve transmissible cancer lineage in M. trossulus itself. Sci Rep 2021;11:1-13. https://doi.org/10.1038/s41598-021-85098-5 |
||||
| 15 Hammel M, Simon A, Arbiol C, Villalba A, Burioli E-V, Pépin JF, Lamy JB, Benabdelmouna A, Bernard I, Houssin M, Charrière GM, Destoumieux-Garzon D, Welch JJ, Metzger MJ, Bierne N: Prevalence and polymorphism of a mussel transmissible cancer in Europe. Mol Ecol 2022;31:736-751. https://doi.org/10.1111/mec.16052 |
||||
| 16 Michnowska A, Hart S, Smolarz K, Hallmann A, Metzger MJ: Horizontal transmission of disseminated neoplasia in the widespread clam Limecola balthica from the Southern Baltic Sea. Mol Ecol 2022;31:3128-3136. https://doi.org/10.1111/mec.16464 |
||||
| 17 Dairain A, Engelsma MY, Drent J, Dekker R, Thieltges DW: High prevalences of disseminated neoplasia in the Baltic tellin Limecola balthica in the Wadden Sea. Dis Aquat Organ 2020;138:89-96. https://doi.org/10.3354/dao03448 |
||||
| 18 Pekkarinen M: Neoplastic Diseases in the Baltic Macoma balthica (Bivalvia) off the Finnish Coast. J Invertebr Pathol 1993;61:138-146. https://doi.org/10.1006/jipa.1993.1026 |
||||
| 19 Thiriot-Quiévreux C, Wolowicz M: Chromosomal study of spatial variation of the prevalence of a gill neoplasia in Macoma balthica (L.) from the Gulf of Gdansk (Baltic Sea). Ophelia 2001;54:75-81. https://doi.org/10.1080/00785326.2001.10409456 |
||||
| 20 Smolarz K, Renault T, Wołowicz M: Ultrastructural study of neoplastic cells in Macoma balthica (Bivalvia) from the Gulf of Gdansk (Poland). J Invertebr Pathol 2006;92:79-84. https://doi.org/10.1016/j.jip.2006.04.003 |
||||
| 21 Weinberg F, Chandel NS: Reactive oxygen species-dependent signaling regulates cancer. Cell Mol Life Sci 2009;66:3663-3673. https://doi.org/10.1007/s00018-009-0099-y |
||||
| 22 Galluzzi L, Kepp O, Heiden MGV, Kroemer G: Metabolic targets for cancer therapy. Nat Rev Drug Discov 2013;12:829-846. https://doi.org/10.1038/nrd4145 |
||||
| 23 Maiuri MC, Kroemer G: Essential role for oxidative phosphorylation in cancer progression. Cell Metab 2015;21:11-12. https://doi.org/10.1016/j.cmet.2014.12.013 |
||||
| 24 Zurburg W, de Zwaan A: The Role of Amino Acids in Anaerobiosis and Osmoregulation in Bivalves. J Exp Zool 1981;215:315-325. https://doi.org/10.1002/jez.1402150309 |
||||
| 25 Kube S, Sokolowski A, Jansen JM, Schiedek D: Seasonal variability of free amino acids in two marine bivalves, Macoma balthica and Mytilus spp., in relation to environmental and physiological factors. Comp Biochem Physiol Part A Mol Integr 2007;147:1015-1027. https://doi.org/10.1016/j.cbpa.2007.03.012 |
||||
| 26 Sokolowski A, Wolowicz M, Hummel H: Free amino acids in the clam Macoma balthica L. (Bivalvia, Mollusca) from brackish waters of the southern Baltic Sea. Comp Biochem Physiol Part A Mol Integr 2003;134:579-592. https://doi.org/10.1016/S1095-6433(02)00360-4 |
||||
| 27 Demas GE, Adamo SA, French SS: Neuroendocrine‐immune crosstalk in vertebrates and invertebrates: implications for host defence. Funct Ecol 2011;25:29-39. https://doi.org/10.1111/j.1365-2435.2010.01738.x |
||||
| 28 Olkowicz M, Debski J, Jablonska P, Dadlez M, Smolenski RT: Application of a new procedure for liquid chromatography/mass spectrometry profiling of plasma amino acid-related metabolites and untargeted shotgun proteomics to identify mechanisms and biomarkers of calcific aortic stenosis. J Chromatogr A 2017;1517:66-78. https://doi.org/10.1016/j.chroma.2017.08.024 |
||||
| 29 Schnaitman C, Greenawalt JW: Enzymatic properties of the inner and outer membranes of rat liver mitochondria. J Cell Biol 1968;38:158-175. https://doi.org/10.1083/jcb.38.1.158 |
||||
| 30 Walker JM, Hammond JBW, Kruger NJ: The Bradford method for protein quantitation. Methods Mol Biol 1988;3:25-32. https://doi.org/10.1385/0-89603-126-8:25 |
||||
| 31 Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem 1951;193:265-275. https://doi.org/10.1016/S0021-9258(19)52451-6 |
||||
| 32 Peterson GL: A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal Biochem 1977;83:346-356. https://doi.org/10.1016/0003-2697(77)90043-4 |
||||
| 33 Habig WH, Pabst MJ, Jakoby WB: Glutathione S-transferases. The first enzymatic step in mercapturic acid formation J Biol Chem 1974;249:7130-7139. https://doi.org/10.1016/S0021-9258(19)42083-8 |
||||
| 34 Jakubowska M, Białowąs M, Stankevičiūtė M, Chomiczewska A, Pažusienė J, Jonko-Sobuś K, Hallmann A, Urban-Malinga B: Effects of chronic exposure to microplastics of different polymer types on early life stages of sea trout Salmo trutta . Sci Total Environ 2020;740:139922. https://doi.org/10.1016/j.scitotenv.2020.139922 |
||||
| 35 Baptista M, Repolho T, Maulvault AL, Lopes VM, Narciso L, Marques A, Bandarra N, Rosa R: Temporal dynamics of amino and fatty acid composition in the razor clam Ensis siliqua (Mollusca: Bivalvia). Helgol Mar Res 2014;68:465-482. https://doi.org/10.1007/s10152-014-0402-7 |
||||
| 36 Chen DW, Su J, Liu XL, Yan DM, Lin Y, Jiang WM, Chen XM: Amino acid profiles of bivalve mollusks from Beibu Gulf, China. J Aquat Food Prod Technol 2012;21:369-379. https://doi.org/10.1080/10498850.2011.604820 |
||||
| 37 Hummel H, Amiard-Triquet C, Bachelet G, Desprez M, Marchand J, Sylvand B, Amiardb JC, Rybarczyk H, Bogaards RH, Sinke J, de Wolf L: Free amino acids as a biochemical indicator of stress in the estuarine bivalve Macoma balthica . Sci Total Environ 1996;188:233-241. https://doi.org/10.1016/0048-9697(96)05177-7 |
||||
| 38 Felbeck H, Wiley S: Free D-amino acids in the tissues of marine bivalves. Biol Bull 1987;173:252-259. https://doi.org/10.2307/1541877 |
||||
| 39 Tabakaeva OV, Tabakaev AV: Amino-Acid Composition of Soft Tissues of the Far-East Bivalve Mollusk Anadara broughtonii . Chem Nat Compd 2016;52:468-471. https://doi.org/10.1007/s10600-016-1674-8 |
||||
| 40 Haider F, Falfushynska HI, Timm S, Sokolova IM: Effects of hypoxia and reoxygenation on intermediary metabolite homeostasis of marine bivalves Mytilus edulis and Crassostrea gigas . Comp. Biochem Physiol A Mol Integr Physiol 2020;242:110657. https://doi.org/10.1016/j.cbpa.2020.110657 |
||||
| 41 Yang L, Venneti S, Nagrath D. Glutaminolysis: A Hallmark of cancer metabolism. Annu Rev Biomed Eng 2017;19:163-94. https://doi.org/10.1146/annurev-bioeng-071516-044546 |
||||
| 42 Jacque N, Ronchetti AM, Larrue C, Meunier G, Birsen R, Willems L, Saland E, Decroocq J, Maciel TT, Lambert M, Poulain L, Hospital MA, Sujobert P, Joseph L, Chapuis N, Lacombe C, Moura IC, Demo S, Sarry JE, Recher C, et al.: Targeting glutaminolysis has antileukemic activity in acute myeloid leukemia and synergizes with BCL-2 inhibition. Blood 2015;126:1346-1356. https://doi.org/10.1182/blood-2015-01-621870 |
||||
| 43 Matre P, Velez J, Jacamo R, Qi Y, Su X, Cai T, Chan SM, Lodi A, Sweeney SR, Ma H, Davis RE, Baran N, Haferlach T, Su X, Flores ER, Gonzalez D, Konoplev S, Samudio I, DiNardo C, Majeti R, et al.: Inhibiting glutaminase in acute myeloid leukemia: metabolic dependency of selected AML subtypes. Oncotarget 2016;7:79722-79735. https://doi.org/10.18632/oncotarget.12944 |
||||
| 44 Willems L, Jacque N, Jacquel A, Neveux N, Maciel TT, Lambert M, Schmitt A, Poulain L, Green AS, Uzunov M, Kosmider O, Radford-Weiss I, Moura IC, Auberger P, Ifrah N, Bardet V, Chapuis N, Lacombe C, Mayeux P, Tamburini J, et al.: Inhibiting glutamine uptake represents an attractive new strategy for treating acute myeloid leukemia. Blood 2013;122:3521-3532. https://doi.org/10.1182/blood-2013-03-493163 |
||||
| 45 Otto-Ślusarczyk D, Graboń W, Mielczarek-Puta M: Aspartate aminotransferase-key enzyme in the human systemic metabolism. Post Hig Med Dosw 2016;70:70. https://doi.org/10.5604/17322693.1197373 |
||||
| 46 Czarnecka AM, Golik P, Bartnik E: Mitochondrial DNA mutations in human neoplasia. J Appl Genet 2006;47:67-78. https://doi.org/10.1007/BF03194602 |
||||
| 47 Sen S, Kawahara B, Chaudhuri G: Mitochondrial-associated nitric oxide synthase activity inhibits cytochrome c oxidase: Implications for breast Cancer. Free Radic Biol Med 2013;57:210-220. https://doi.org/10.1016/j.freeradbiomed.2012.10.545 |
||||
| 48 Ottaviani E, Franchini A, Franceschi C: Presence of several cytokine-like molecules in molluscan hemocytes. Biochem Biophys Res Commun 1993;195:984-988. https://doi.org/10.1006/bbrc.1993.2141 |
||||
| 49 Cooper EL, Arizza V, Cammarata M, Pellerito L, Parrinello N: Tributyltin affects phagocytic activity of Ciona intestinalis hemocytes. Comp Biochem Physiol C 1995;112:285-289. https://doi.org/10.1016/0742-8413(95)02022-5 |
||||
| 50 Babior BM: The respiratory burst oxidase. Adv Enzymol Relat Areas Mol Biol 1992;65:49-95. https://doi.org/10.1002/9780470123119.ch2 |
||||
| 51 Segal AW, Shatwell KP: The NADPH oxidase of phagocytic leukocytes. Ann N Y Acad Sci 1997;832:215-222. https://doi.org/10.1111/j.1749-6632.1997.tb46249.x |
||||
| 52 Cui S, Reichner JS, Mateo RB, Albina JE: Activated murine macrophages induce apoptosis in tumor cells through nitric oxide-dependent or -independent mechanisms. Cancer Res 1994;54:2462-2467. | ||||
| 53 Adamo SA: Comparative psychoneuroimmunology: Evidence from the insects. Behav Cogn Neurosci Rev 2006;5:128-140. https://doi.org/10.1177/1534582306289580 |
||||
| 54 Boland R, Joyce BJ, Wallace MJ, Stanton H, Fosang AJ, Pierce RA, Harding R, Hooper SB: Cortisol enhances structural maturation of the hypoplastic fetal lung in sheep. Physiol J 2004; 554:505-517. https://doi.org/10.1113/jphysiol.2003.055111 |
||||
| 55 Binder ARD, Pfaffl MW, Hiltwein F, JuergenGeis J, Beggel S: Does environmental stress affect cortisol biodistribution in freshwatermussels? Conserv Physiol 2019;7:coz101. https://doi.org/10.1093/conphys/coz101 |
||||
| 56 Lupo C, Prisco DI, Dessi' Fulgheri F, Tomasucci M: Identification and biosynthesis of steroids in the marine molluscs Aplysia Depilans. Comp Biochem Physiol B Biochem Mol 1973;45:303-310. https://doi.org/10.1016/0305-0491(73)90065-5 |
||||
| 57 Lee SR, Kim HK, Youm JB, Dizon LA, Song IS, Jeong SH, Seo DY, Ko KS, Rhee BD, Kim N, Hanet J: Non-genomic effect of glucocorticoids on cardiovascular system. Pflugers Arch 2012;464:549-559. https://doi.org/10.1007/s00424-012-1155-2 |
||||