×
![]()
Corresponding Author: Shibashish Giri
Center for Biotechnology and Biomedicine, Department of Cell Techniques and Applied Stem Cell Biology, Medical Faculty of University of Leipzig, Deutscher Platz 5, D-04103 Leipzig (Germany)
Tel. +49-3419731353, Fax +49-3419731329, E-Mail shibashish.giri@bbz.uni-leipzig.de; shibashishgermany@gmail.com
Improved Metabolic Pathways of Glycolysis, Glycogen Synthesis, the Urea Cycle, and Cytochrome Peroxidase Oxidative Reabsorption in a Miniature Bioreactor
Shibashish Giria,b Wolfgang Schmidt-Heckc Augustinus Badera
aCenter for Biotechnology and Biomedicine, Department of Cell Techniques and Applied Stem Cell Biology, University of Leipzig, Leipzig, Germany, bDepartment of Plastic and Hand Surgery, University Hospital Rechts der Isar, Munich Technical University, Munich, Germany, cDepartment of Molecular and Applied Microbiology, Leibniz Institute for Natural Product Research and Infection, Hans Knöll-Institute, Jena, Germany
Introduction
There is increasing clinical interest in the development of bioartificial liver support systems, which require adequate number of functional hepatocytes because these cells carry out most hepatic metabolic functions [1]. It has been reported that hepatocytes rapidly lose their metabolic and biosynthetic capabilities in conventional cultures and standard in vitro culture conditions [2]. Hence, long-term functional maintenance of primary adult hepatocytes is a major challenge for the development of bioartificial liver support, which acts as a temporary liver support for patients suffering from liver failure or as a bridge to liver transplantation or regeneration [3]. Spontaneous liver regeneration among acute liver patients occurs in approximately 40% of acute failure cases [3]. Although there are many factors crucial for the long-term culture of adult hepatocytes, oxygen is a very important nutrient for numerous hepatocyte functions, and it is a very critical factor for the optimization of hepatocyte microenvironment in vitro [4]. Kidambi et al. reported that a 95% oxygen atmosphere improves hepatic functions [5], while other researchers have reported that high oxygen and partial pressure creates reactive oxygen pressure and causes oxidative damage to hepatocytes [6]. Recently, Xiao et al. reported that the expression of 84 drug metabolism-related genes was the closest to physiological levels in a sandwich culture model [7]. Therefore, there is an ongoing debate about hepatocyte cultures, either for preclinical research or bioartificial liver support in a clinical setting. In order to successfully develop a bioartificial liver support system, it is necessary to optimize the cellular microenvironment, where oxygen is one of the most important parameters for hepatocyte function. These cells require sufficient oxygen to carry out metabolic functions [8-9] and consume oxygen at a rate 10 times higher than other cell types [10]. Oxygen is supplied to hepatocytes in vivo via a mixture of arterial and venous blood [9]. It is estimated that the oxygen consumption of hepatocytes is higher in vivo than in vitro [11, 12]. Current conventional culture methods rarely focus on enhanced direct oxygenation for adult hepatocytes, as seen in vivo [13]. Several attempts have been made for direct oxygenation in long-term cultures by using different bioreactor devices, because the performance of long-term cultures of primary hepatocytes depends on these bioreactor devices [14–17]. None of the existing bioreactor devices have been monitored, and none have established the metabolic pathways of genes of glycolysis, glycogen synthesis, the urea cycle, cytochrome peroxidase oxidative reabsorption, and UDP-glucuronosyltransferase (UGT). Any abnormality in these pathways negatively influences hepatocyte functions in the bioartificial liver device. For example, a gene mutation for the enzyme UGT has been shown to result in a condition called Gilbert’s syndrome and jaundice [18].
Materials and Methods
Hepatocyte isolation and culture
Hepatocytes were isolated from 12-week-old mice using the two-step method described by Seglen (1976), with slight modifications [19]. Briefly, the liver was pre-perfused in situ with cold Ringer buffer. Connective tissue was digested by perfusing the liver with 0.05% type IV collagenase. Viability of the isolated hepatocytes was more than 90% as assessed by trypan blue exclusion assay. Cells were suspended in Williams E medium supplemented with fetal calf serum (5%), insulin (0.17 U/mL), prednisolone (0.85 mg/mL), glucagon (0.015 mg/mL), glutamine (8.9 mM), penicillin (450 U/mL), and streptomycin (450 mg/mL). The gas-permeable PTFE membrane in the mini bioreactor was moistened with rat tail collagen type I, which was prepared according to the method described by Elsdale and Bard (1972). Cells were seeded at a density of 2.5 × 105 cells/well in the bioreactor or in the plates. After 48 h of incubation at 37 °C in a humidified atmosphere containing 5% CO2 and 20% O2 (v/v), a second collagen gel layer was added to the cells as a modification to the methods of Dunn et al. (1989) and Bader et al. (1992). Hepatocytes were cultured between two collagen layers in a sandwich model (3D configuration) under direct oxygenation (5%, 10%, 21%, and 40% O2). for 14 days. We seeded 2.5 × 105 cells/well and changed the medium once every 24 h. The supernatant was removed from each well every day and stored at −80 °C until further analysis.
Bioreactor configuration: 24-well bioreactor
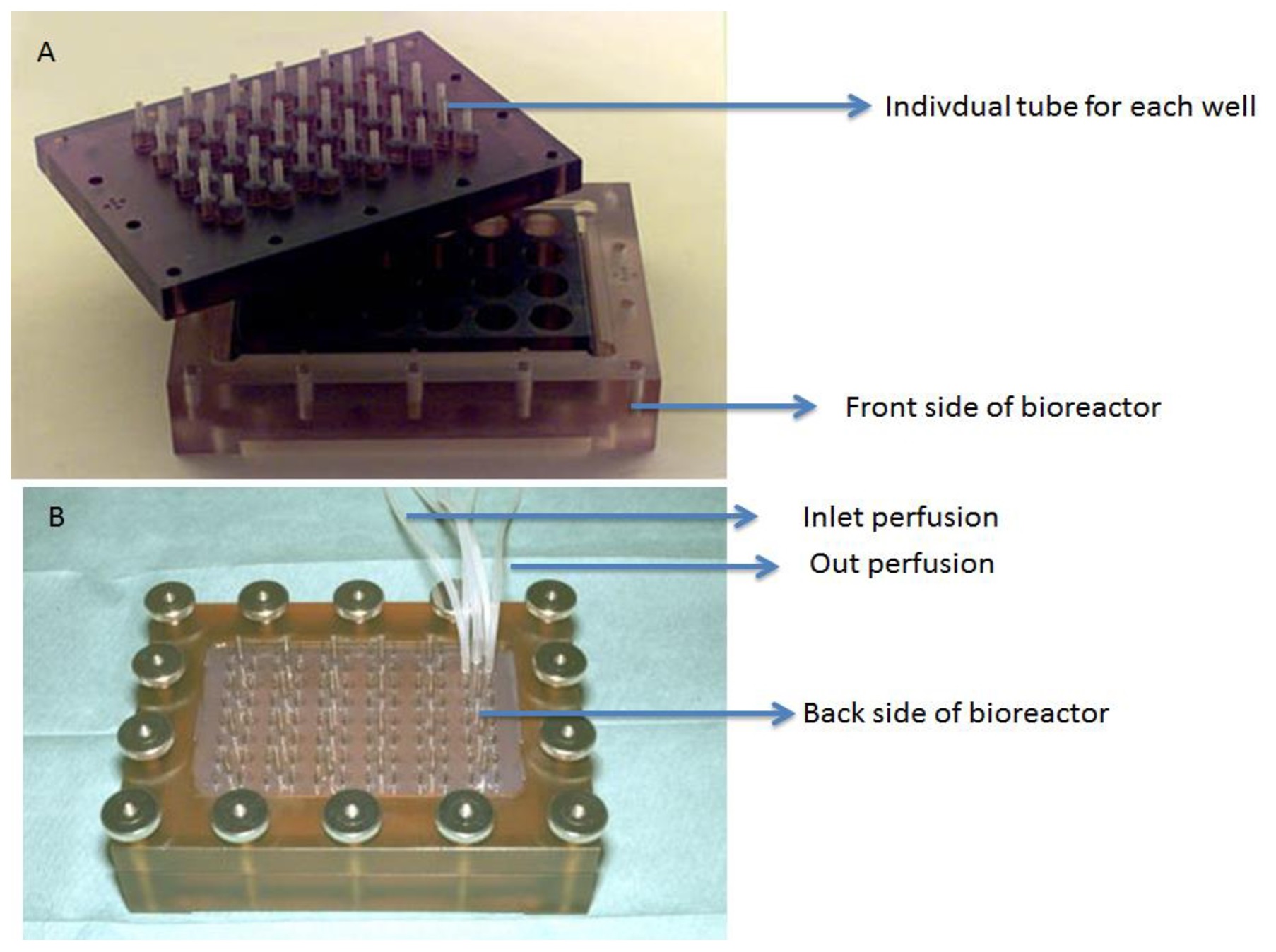
The bioreactor was set up as described in a previous study [20–22]. Briefly, it uses a modified form of the conventional 24-well cell cultivation plate and is composed of a scaffold of polycarbonate with 24 wells, a gas-permeable PTFE membrane of 25 mm thickness, a 24-hole silicon seal, and a 24-hole metal base (Fig. 1). The PTFE membrane is a transparent, thermoplastic film with an oxygen permeability of 114.5 cm3 m−2 24 h−1 k Pa−1 and allows a maximal oxygen supply of 90 mmol per well (1.77 cm2) per day for cell cultures. Based on this model, the amount of oxygen required by the hepatocytes (2.5 × 105 cells/well) in the bioreactor was suggested to range between 6.5 and 19.5 mmol, allowing a 5-14 times higher oxygen supply than needed. The bioreactor was placed into a support that facilitated direct delivery of oxygen to the cells from the bottom of the device.
The miniature bioreactor represents a scaledown of a previously developed clinically flat membrane bioreactor [23-29] to a conventional 24-well cell cultivation plate format. Earlier preclinical tests were performed in a pig model, but did not evaluate the metabolic functions of the genes of the hepatic metabolic pathways, such as glycolysis (glucokinase, phosphofructokinase, and pyruvate kinase), glycogen synthesis (glycogen synthetase, UTP glucose-1-phosphate uridylylisomerase, phosphoglucomutase, and glycogen phosphorylase), the urea cycle (arginase, ornithine carbomoyltransferase, and fumarate hydratase), oxidative reabsorption (peroxidase), and cytochrome peroxides (catalase and SOD) under direct oxygenation conditions since oxygenation is an open question for the hepatocyte culture model. The bioreactor consisted of a polycarbonate scaffold with 24 wells, a 24-hole silicon seal, and a 24-hole metal base to form a scalable 24-well miniature bioreactor. Isolated hepatocytes were cultured between two layers of collagen termed as the organotypical sandwich model. The hepatocytes were located at a distance of about 20 μm in extracellular matrix different from the distance (over 200 μm) in conventional culture conditions. The medium and cells in the miniature bioreactor were oxygenated in the incubator by diffusion of humidified air across non-porous PTFE membranes (25 μm thick). The PTFE membrane is highly permeable to oxygen, allowing enhanced oxygenation from both sides, and acts as an internal oxygenation membrane. This design provides direct oxygenation of the medium and cells in each cultured well by diffusion of humidified air into the incubator. A microporous membrane that separates the cell compartment from the perfusion medium compartment is generally used when the miniature bioreactor is used in continuous flow to protect cells from shear forces and to control mass transfer of different solutes. The flow rate (9 mL/h) in the inlet and outlet streams is supplied by each well through separate silicone tubes. Each cultured cell has its own supply of perfusion medium. Hence, the inlet and outlet remove metabolic waste and supply fresh nutrients in the form of medium. The oxygen-permeable membrane with rat tail type I collagen: after seeding and attachment of the hepatocytes, a second layer of matrix was placed on top as described earlier to establish the organotypical sandwich model. The hydrophilic sides of the membranes are in the inner compartment of the bioreactor and the hydrophobic sides are in the outer compartment. The entirety of the contact surfaces were sealed with silicone adhesive and sterile air filters were connected to regulate air pressure. After 24–48 h, the bioreactor was rinsed with 500 μL of distilled water per well to remove dirt particles and solvent vapors of the glue. The prepared miniature bioreactor was light and dust-protected. The bioreactor was autoclaved for 12 min at 121°C 24–48 h before testing. We tested for leakages by simple injection of sterile water before experimentation.
Semi-quantitative analysis of genes
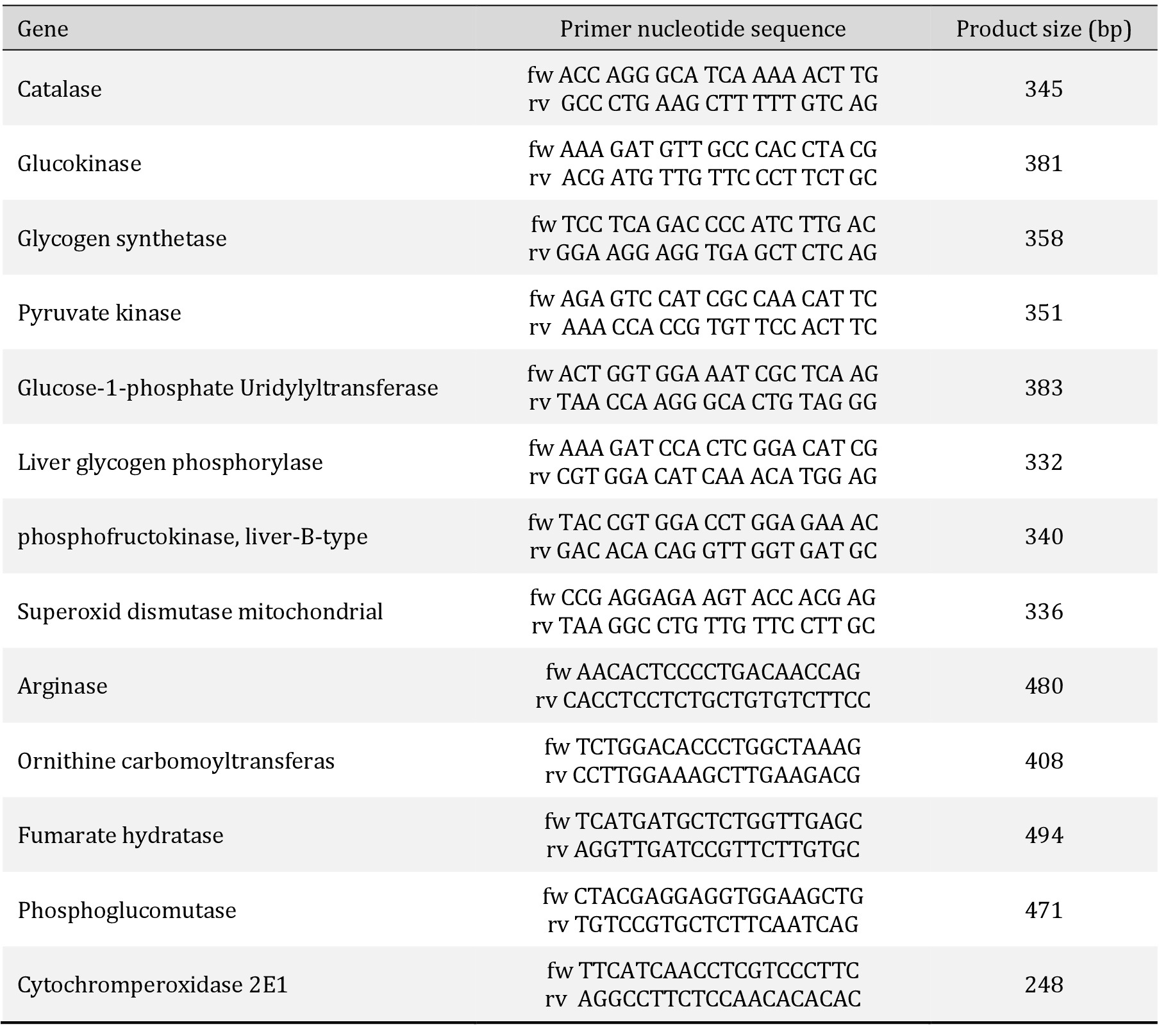
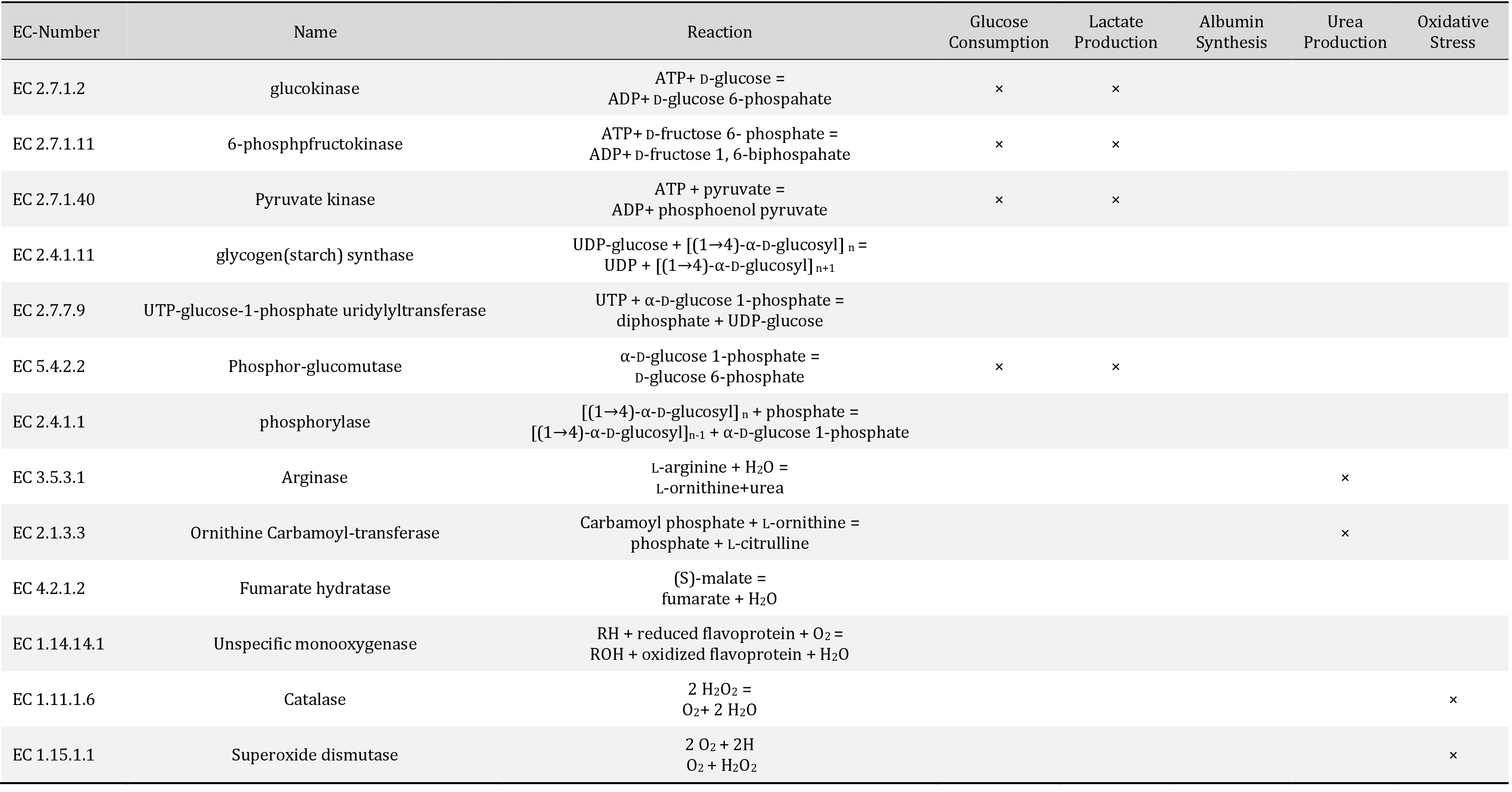
Total RNA of cells was extracted using an RNeasy Mini kit. RT-PCR was performed with OneStep RT-PCR kit on a GeneAmp PCR system 9700 (Applied Biosystems, Foster City, CA) according to the manufacturer’s instructions. The expression of various hepatic-specific genes was analyzed. The sequences of the primary pairs, product sizes, and numbers of PCR cycles are listed in Table 1. Relative expression levels were calculated by measuring the signal intensity. Gene expression data for each gene was normalized to the expression level of glyceraldehyde 3-phosphate dehydrogenase, which was used as an internal control. The expression level of genes in the liver tissue (in vivo) was set as 1 and the expression levels of genes in primary hepatocytes cultured under hypoxia, normoxia, or hyperoxia were compared with those in the in vivo condition. Assignment of EC numbers to metabolic functions is shown in Table 2.
Statistical analysis
All acquired data were statistically analyzed by Student’s t -test. Probability values lower than 0.05 were considered to be statistically significant.
Results
Oxygen plays a significant role in disease development and maintaining normal physiology. Hepatocytes are both consumers and producers of glucose, depending on local conditions. During fasting, hepatocytes have the ability to release glucose into the systematic circulation via the process of glycogenolysis in order to fulfill the energy requirements of the brain, skeletal muscles, and red blood cells. Hepatocytes respond differently to a wide range of oxygenation levels. In the absence of oxygen, the cells lose their plasma membrane integrity, leading to necrotic cell death [29]. Higher oxygenation levels may also alter hepatic functions. Hence, optimal oxygenation of an in vitro culture model is essential for toxicity studies, bioartificial support experiments, and others to compare levels in vivo . We investigated the effect of hypoxia, normoxia, and hyperoxia on hepatocytes and their metabolic activities in a three-dimensional culture model. The metabolic activities of hepatocytes were evaluated based on lactate formation, glucose consumption, albumin secretion, and urea production. Cellular membrane integrity was estimated by measurement of released LDH. A small 24-well bioreactor with direct oxygenation was used to allow optimal oxygen supply for the primary hepatocyte culture. To ensure optimal metabolic functionality of the cultured primary hepatocytes, it is important to provide the cells with an in vitro culture milieu that mimics the physiological environment of the cells. In vivo oxygen concentrations were reconstructed and observed via the reversal of anaerobic metabolic states in vitro . Primary hepatocytes were cultured under optimal oxygen conditions by the use of integrated oxygen-permeable membranes. Overlaying the culture with additional layers of complexity, hepatocellular functions, growth factor response, and hepatocellular response to xenobiotic exposure were modeled.
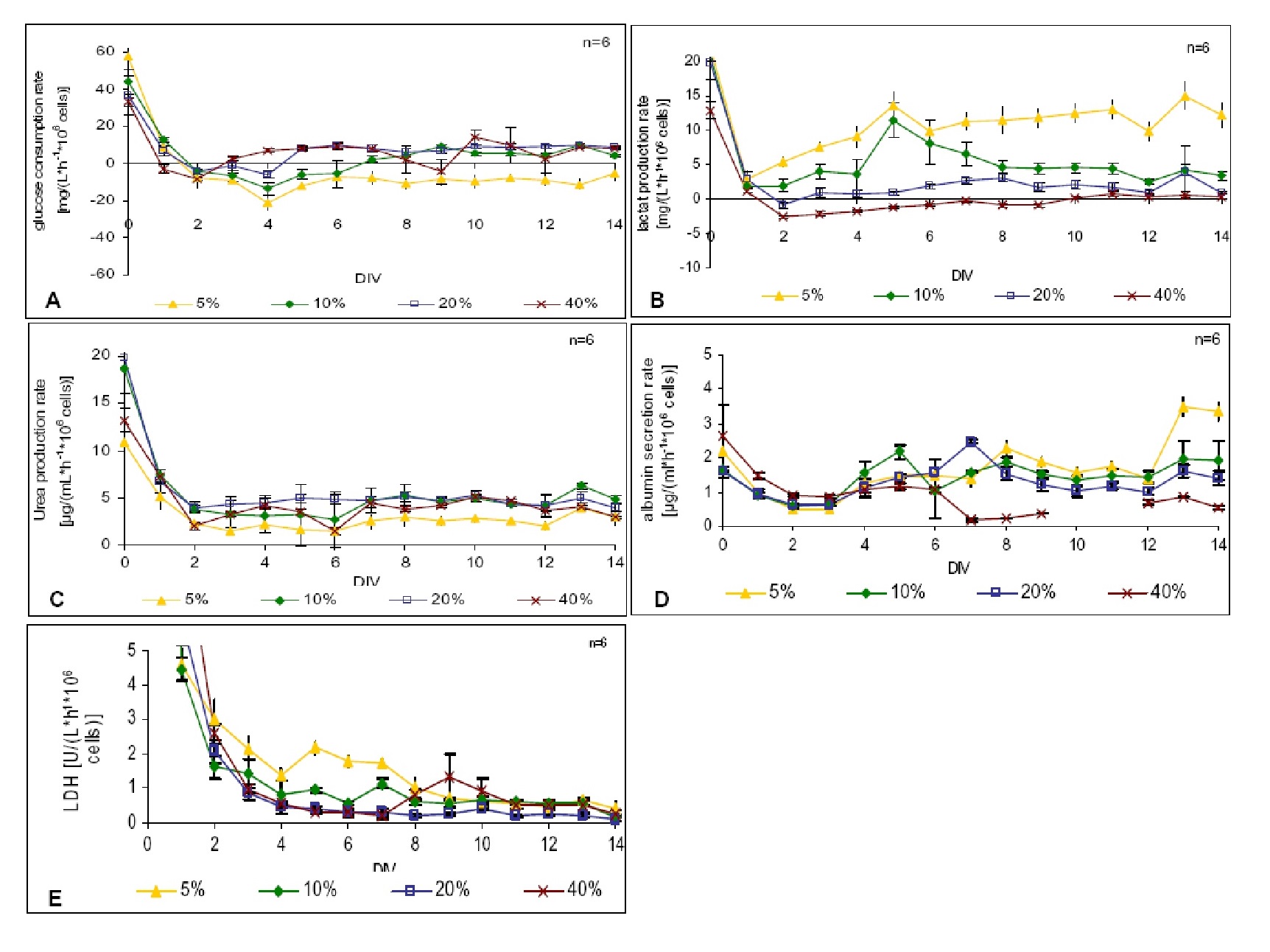
We investigated the effects of hyperoxia, normoxia, and hypoxia on cultured primary adult hepatocytes. Hepatocytes cultured in 10%, 20%, and 40% oxygenation formed a confluent layer within 3 days. In contrast, hepatocytes cultured in 5% O2 could not grow to confluency. Morphological sections of cells cultured under hypoxia (10% O2) showed the presence of intracellular inclusions (granules), indicating that a hypoxic environment has a negative impact on hepatocytes and a 5% O2 supply is insufficient for their growth. A majority of the cells died within the first few days, as demonstrated by increased LDH release. LDH level was high in 5%, 10%, and even 40% oxygenation conditions, but negligible in 20% oxygenation. This indicated that 20% O2 condition provides a good environment for cell growth from day 1 to day 14. A majority of cells died within the first few days, as demonstrated by increased LDH release (Fig. 2). The presence of intracellular inclusions (granules) indicated that the hypoxic environment has a negative impact on hepatocytes (Fig. 2). Glucose consumption by hepatocytes cultured under hypoxia (5% and 10% O2) was enhanced (Fig. 3A). The increased rate of urea secretion by hepatocytes cultured under normoxia and hyperoxia conditions within the experimental period (20% and 40% O2) indicated optimal metabolic activity (Fig. 3C). Albumin secretion was found to be less in cells under hyperoxia (Fig. 3D) than that in cells under hypoxia (5% and 10%). This may be attributed to our bioreactor, based on direct oxygenation via PTFE membrane in a 3D configuration, which may promote the albumin secretion function of cells cultured even in low O2 conditions. To assess the contribution of glycolysis in energy formation in liver cells, the rate of lactate formation was evaluated. Lactate production was very high in 5% and 10% oxygenation conditions, but negligible in 20% oxygenation. Almost no lactate formation was observed in 40% oxygenation condition (Fig. 3B).
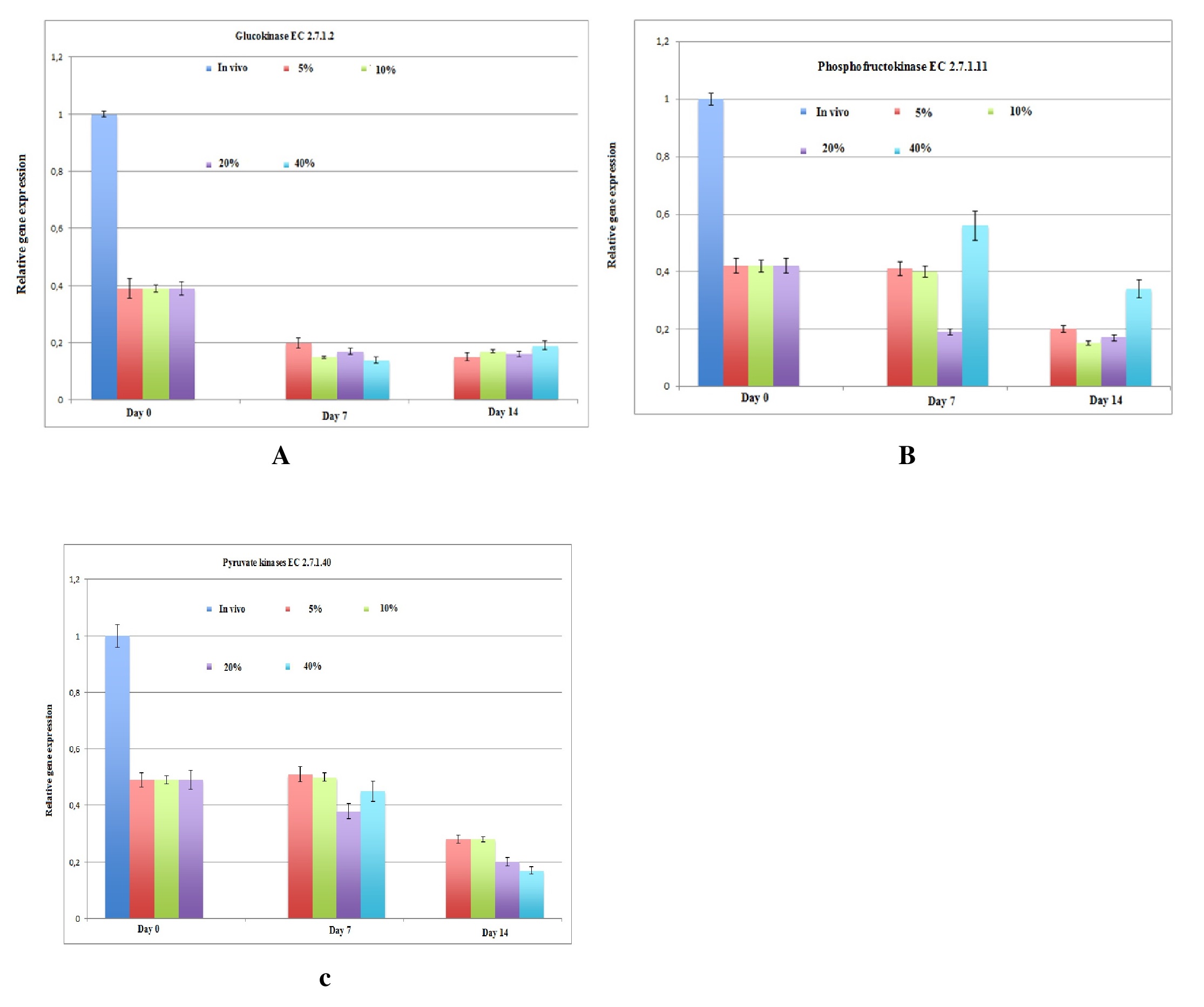
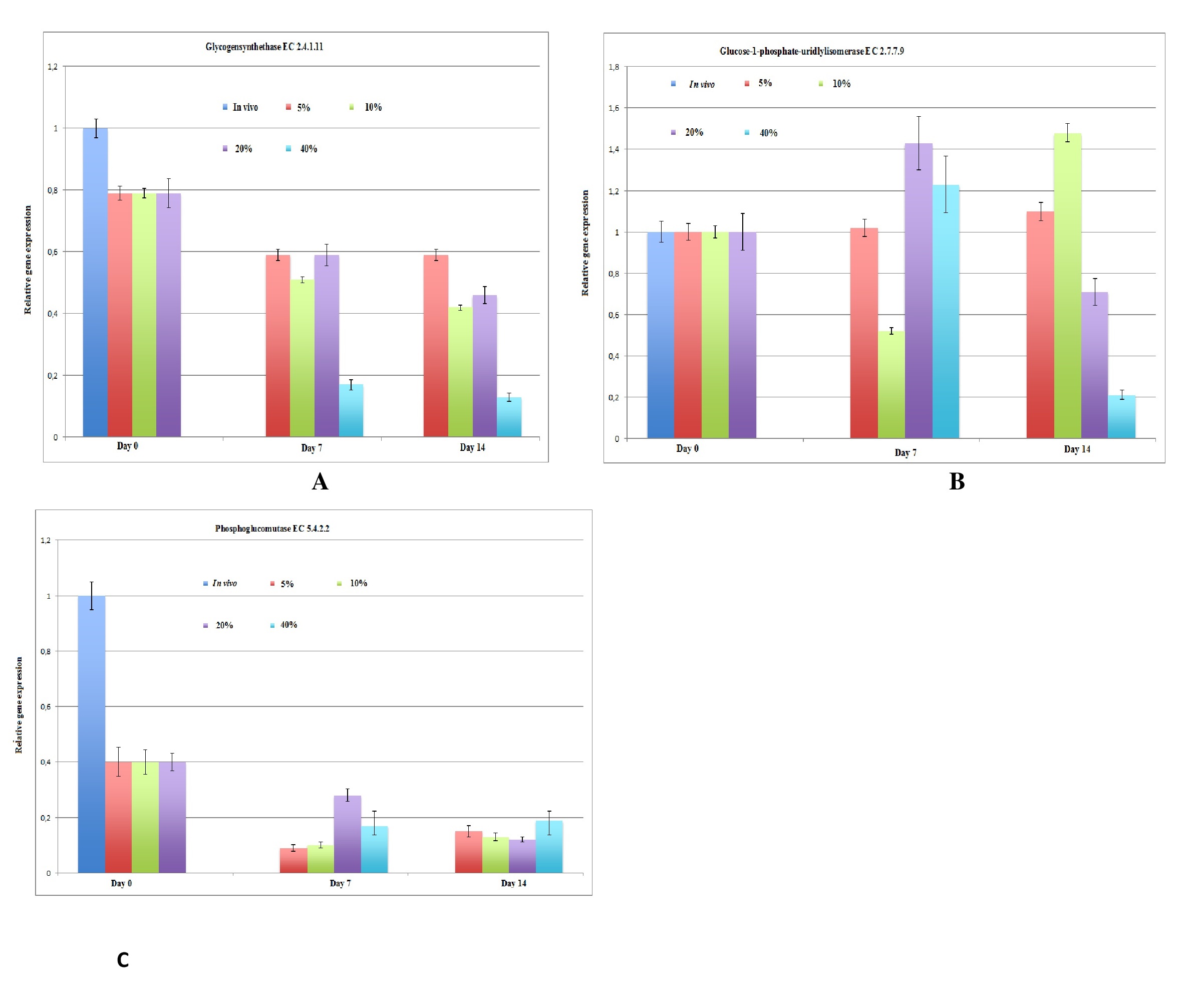
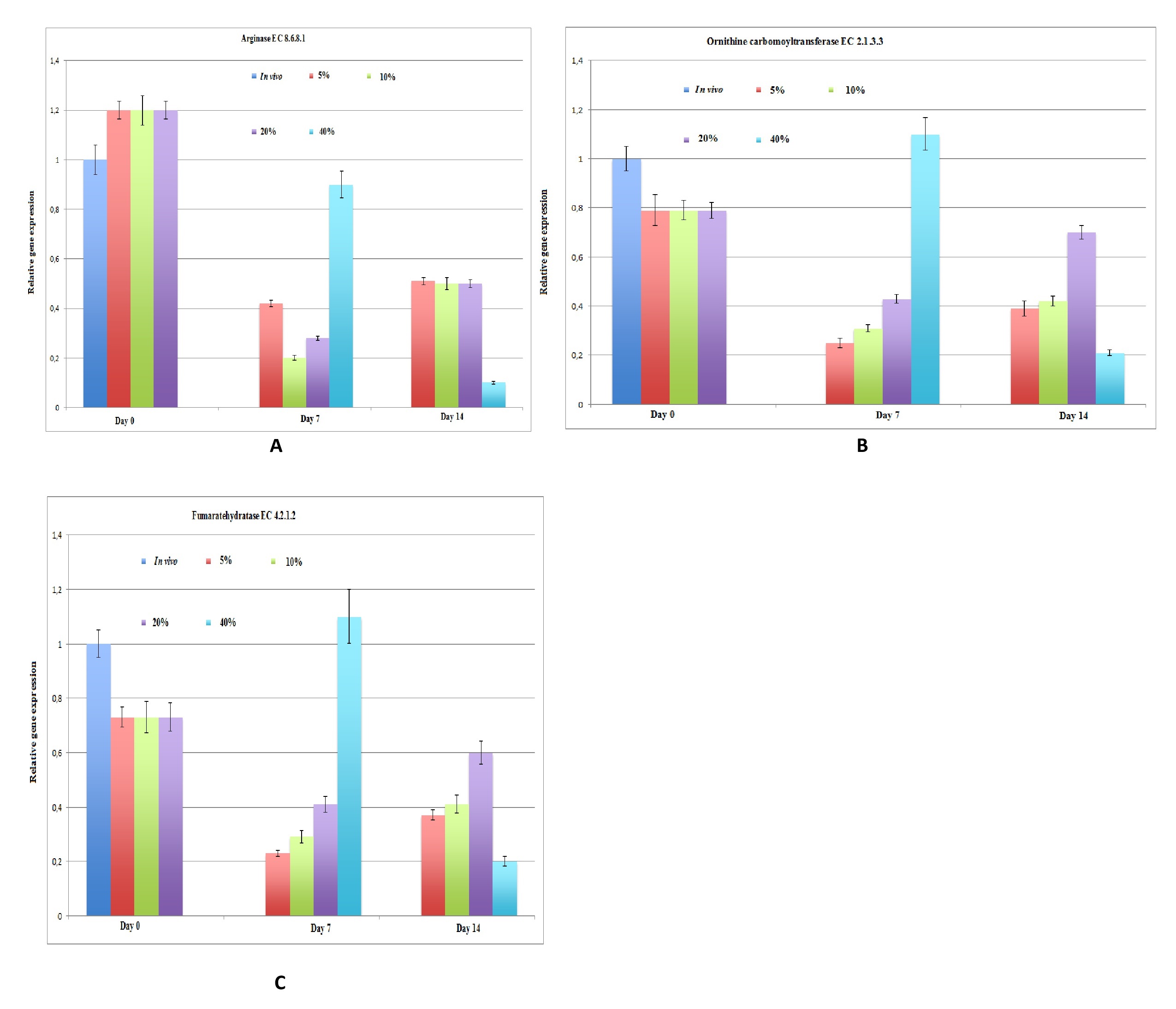
Additionally, the expression of various hepatic-specific genes in hepatocytes cultured under various oxygen levels was determined using semi-quantitative PCR. Although gluconeogenesis in vivo has been studied by biochemists and physiologists, there are little data for glycolysis and gluconeogenesis in in vitro long-term cultures via direct oxygenation in a 3D configuration for up to 14 days. We evaluated the mRNA expression of glycolysis enzymes under different oxygen conditions and compared it with the in vivo mRNA expression levels of the glycolysis enzymes. The levels of glucokinase and phosphofructokinase were downregulated in all hepatic cultures under different oxygenation conditions (Fig. 4A, B). The expression of the other glycolysis enzyme, pyruvate kinase, in hypoxic cultures corresponded to the metabolic results for glucose consumption and lactate formation (Fig. 4C). We isolated individual pathways of these glycolysis enzymes in metabolic mini-maps of glycolysis designed by Donald Nicholson (International Union of Biochemistry and Molecular Biology). We also analyzed the process of glycogen synthesis by which glucose is stored in the body in the form of glycogen in the liver and muscles. Due to low concentration of glycogen in the muscles, only the glycogen stored in the liver is accessible to other organs that can be quickly mobilized to meet a sudden need for glucose. The expression levels of genes encoding enzymes for glycogen degradation were found to be downregulated (Fig. 5C), while those encoding enzymes for glycogen synthesis were expressed at a normal level (Fig. 5A, B) in all cases. In addition to glycolysis and glycogen synthesis, the liver detoxifies ammonia through the urea cycle. Gene expression levels of enzymes involved in urea synthesis are shown in Fig. 6. The expression of arginase, ornithine carbomoyltransferase, and fumarate hydratase was upregulated at day 7 under all oxygen conditions, and downregulated at day 14 under 40% oxygenation.
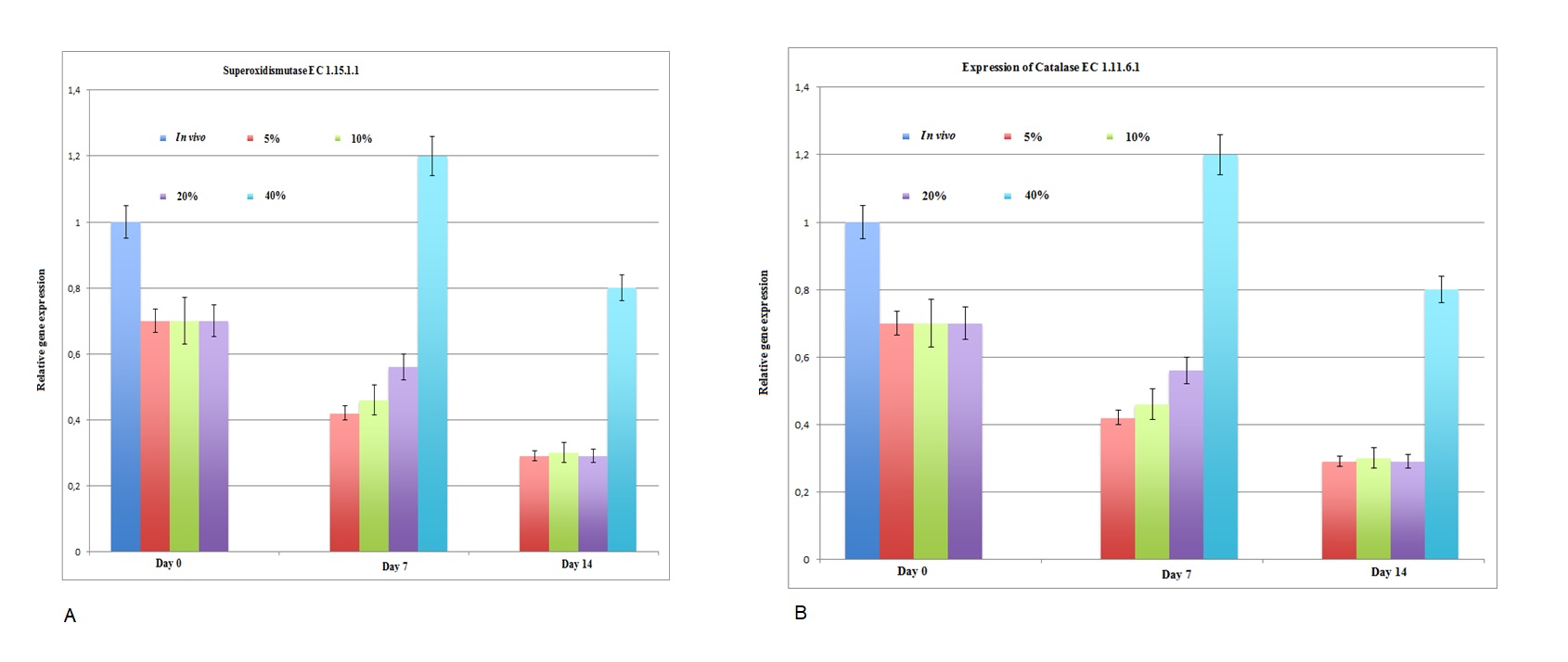
Apart from glycolysis, glycogen synthesis, and urea cycle analysis, the toxic profile of a few free radicals, such as superoxide and hydroxide, and their response to adult hepatocytes in different oxygenation conditions are very important parameters for the development of bioreactor-based bioartificial liver support. Toxic molecules such as superoxide and hydroxide radicals can be found in cells due to the presence of oxygen along with other products of normal metabolic processes. Superoxide is the most important free radical in the body. SOD is also responsible for the conversion of superoxide into oxygen and peroxide. Catalase is frequently used by cells to rapidly catalyze the decomposition of hydrogen peroxide into less reactive gaseous oxygen and water molecules. The effect of hyperoxia (40% O2) on hepatocytes is shown in Fig. 7, where the mRNA expression of catalase and SOD was upregulated (Fig. 7). Hydrogen peroxide is destroyed by peroxidase, which reduces H2O2 to water. The biotransformation activity was only presented by the expression of the cytochrome peroxidase gene (CYP 2E1 gene), which was downregulated in all cases (Fig. 8A). In addition to these free radicals, we analyzed the expression of the liver-specific enzyme UGT, which is responsible for bilirubin disposal. An abnormality in this enzyme results in Gilbert’s syndrome and jaundice. The expression of phase II enzyme UGT is upregulated in 20% oxygen condition compared with that in other oxygen conditions (Fig. 8B). Taken together, 20% oxygenation is an optimal condition for almost all experiments in this paper for the development of bioreactor-based bioartificial liver support. The optimum oxygenation for albumin production, urea synthesis, glucose consumption, and lactate production is shown in Supplementary Fig. 1A. The assignment of EC numbers to Kyoto Encyclopedia of Genes and Genomes pathway is shown in Supplementary Fig. 1B. The metabolic pathways of glycolysis, the urea cycle, glycogen synthesis, and galactose metabolism are shown in Supplementary Fig. 1B–E, respectively (for all supplementary material see www.cellphysiolbiochem.com).

Discussion
Generally, oxygen is depleted very quickly compared with other nutrients (glucose and amino acids). Proper oxygenation is important to many aspects of the liver phenotype so that the cultured hepatocytes do not become hypoxic and can meet all metabolic demands. The effects of growth factors, peptide hormones, and cytokines are greatly influenced and greatly disrupted by high perfusion rates in vitro. Previously, we evaluated hepatic functions in our bioreactor device. We created a highly tailored microenvironment. Our bioreactor devices aimed towards large-scale culture for extracorporeal liver support as well as drug development. Perfusion culture conditions are superior for metabolic functions such as urea genesis compared with static culture conditions [30], while insulin uptake and stimulation of glucokinase and pyruvate kinase are more physiological in the perfused system [31]. The perfusion flow of the present bioreactor device was established previously in preclinical research and in a pig model in a translational setting. There was significant improvement compared to previous bioreactor devices, particularly for evaluating the potential effects of oxygenation on the genes of hepatic metabolic pathways, such as glycolysis (glucokinase, phosphofructokinase, and pyruvate kinase), glycogen synthesis (glycogen synthetase, UTP glucose-1-phosphate uridylylisomerase, phosphoglucomutase, and glycogen phosphorylase), the urea cycle (arginase, ornithine carbomoyltransferase, fumarate hydratase), oxidative reabsorption (peroxidase), and cytochrome peroxides (catalase and SOD), which might have implications in liver tissue engineering as well as bioartificial liver support, since our bioreactor simulates the liver microenvironment of liver.
Catalase is an enzyme that protects the hepatocytes against oxidative injury [32] by breaking down toxic hydrogen peroxide to harmless water and oxygen. Accumulation of hydrogen peroxide is dangerous and causes hepatocyte damage and liver cirrhosis. Our bioreactor device could be an alternative for temporary detoxification and synthesis functions of the liver. Ammonia accumulation is a common feature in acute liver failure patients. It has been reported that accumulated or increased ammonia negatively affects spontaneous liver regeneration [33. 34]. The urea cycle is a phenomenon in which waste (ammonia) is removed from the body. The result of liver failure is accumulation of nitrogenous waste, mainly ammonia, which leads to cerebral edema and causes intracranial pressure. Intracranial hypertension remains an unsolved clinical problem and a frequent cause of death among fulminant liver failure patients. Interestingly, our bioreactor device controlled intracranial pressure in a pig model and improved survival benefits. The average waiting time for the most urgent listing for acute liver failure patients is 6 days according to United States National Organ Sharing data. Sometimes, several patients have urgently needed transplantation within 1–2 days [35]. Unlike liver dialysis, which provides only detoxification of blood toxins from the patient’s blood, this presented device can provide synthetic detoxification, and thus might serve as a better liver support for spontaneous liver regeneration or act as a bridge until transplantation. The desired aim of our proposed device is to provide short-term (a few days to some weeks) metabolic and temporary liver support for liver failure patients until a suitable donor organ becomes available or spontaneous liver regeneration occurs.
The bioreactor demonstrated in this study successfully improved glucose metabolism, urea genesis, gluconeogenesis, oxidative reabsorption, cytochrome peroxides, and hepatic functions. Additionally, these successful optimizations confirm the feasibility of this highly tailored bioreactor device for liver organ-on-a-chip experimentation and as an extracorporeal liver support system. Furthermore, application of human induced pluripotent stem cell-derived hepatocytes in this device is promising, but requires further study.
Conclusion
We presented miniaturization of liver tissue analogues for improved in vitro performance of liver functions, similar to liver behavior in vivo . We evaluated the metabolic pathways of glycolysis, glycogen synthesis, the urea cycle, and cytochrome peroxidase oxidative reabsorption as well as essential hepatic functions in a clinically relevant module for the generation of a biomimetic in vivo liver microenvironment. An abnormality in these pathways has a negative influence on the cultured hepatocytes. After evaluation, we found that 20% oxygenation was optimum for long-term culture of hepatocytes, and may serve as an alternative approach for hepatic tissue engineering. Reduced oxygen delivery to the liver is a common and important clinical problem for surgeons and for bioartificial liver support-oriented hepatocyte research. Hepatocytes may encounter stress from oxygen deprivation in various conditions, like fasting or toxicity. It is crucial to develop effective in vitro techniques with optimal oxygen conditions, that are suitable for the long-term survival of hepatocytes cultures at high densities, which, along with functional maintenance, may contribute to the establishment of bioreactor-based bioartificial liver support systems. The use of functional hepatocytes cultured in small-scale bioreactors with 20% oxygen for long-term culture may be an alternative approach to overcome the limitations of conventional as well as standard in vitro culture conditions for bioartificial liver support. Considering the simplicity of our scalable design and improvement of major liver functions in this miniature bioreactor system, we envision that this methodological approach could potentially be used for bioartificial liver construction.
The authors would like to thank Claudia Pösel for technical assistance and Prof. Grilles for valuable suggestions.
Author Contributions
Performance of experiments and analysis of data were done by Shibashish Giri. Augustinus Bader has supervised and discussed this project. Wolfgang Schmidt-Heck generated the idea about analysis of the data of gene expression and performed.
Funding
This study was supported by the Hepatocyte Endocytosis Project.
The authors declare that no conflicts of interest exist.
| 1 Struecker B, Raschzok N, Sauer IM: Liver support strategies: cutting-edge technologies. Nat Rev Gastroenterol Hepatol 2014;11:166-176. https://doi.org/10.1038/nrgastro.2013.204 |
||||
| 2 LeCluyse EL, Bullock P, Parkinson A: Strategies for restoration, maintenance of normal hepatic structure and function in long term culture of rat hepatocytes. Adv Drug Deliv Rev 1996;22:133-186. https://doi.org/10.1016/S0169-409X(96)00418-8 |
||||
| 3 Riordan SM, Williams R: Treatment of hepatic encephalopathy. N Engl J Med 1997;337:473-479. https://doi.org/10.1056/NEJM199708143370707 |
||||
| 4 Jungermann K, Kietzmann T: Oxygen: modulator of metabolic zonation and disease of the liver. Hepatology 2000;31:255-260. https://doi.org/10.1002/hep.510310201 |
||||
| 5 Kidambi S, Yarmush RS, Novik E, Chao P, Yarmush ML, Nahmias Y: Oxygen-mediated enhancement of primary hepatocyte metabolism, functional polarization, gene expression, and drug clearance. Proc Natl Acad Sci U S A 2009;106:15714-15719. https://doi.org/10.1073/pnas.0906820106 |
||||
| 6 Yan HM, Ramachandran A, Bajt ML, Lemasters JJ, Jaeschke H: The oxygen tension modulates acetaminophen-induced mitochondrial oxidant stress and cell injury in cultured hepatocytes. Toxicol Sci 2010;117:515-523. https://doi.org/10.1093/toxsci/kfq208 |
||||
| 7 Xiao W, Shinohara M, Komori K, Sakai Y, Matsui H, Osada T: The importance of physiological oxygen concentrations in the sandwich cultures of rat hepatocytes on gas-permeable membranes. Biotechnol Prog 2014;30:1401-1410. https://doi.org/10.1002/btpr.1954 |
||||
| 8 Lemasters JJ, Ji S, Thurman RG: Centrilobular injury following hypoxia in an isolated, perfused rat liver. Science 1981;213:661-663. https://doi.org/10.1126/science.7256265 |
||||
| 9 Lemasters JJ: Hypoxic, ischemic, and reperfusion injury to liver, in Arias IM (ed): The Liver: Biology and Pathobiology. Philadelphia, Lippincott Williams & Wilkins, 2001, ed 4, pp. 257-279. | ||||
| 10 Smith MD, Smirthwaite AD, Cairns DE, Cousins RB, Gaylor JD: Techniques for measurement of oxygen consumption rates of hepatocytes during attachment and post-attachment. Int J Artif Organs 1996;19:36-44. https://doi.org/10.1177/039139889601900106 |
||||
| 11 Balis UJ, Behnia K, Dwarakanath B, Bhatia SN, Sullivan SJ, Yarmush ML, Toner M: Oxygen consumption characteristics of porcine hepatocytes. Metab Eng 1999;1:49-62. https://doi.org/10.1006/mben.1998.0105 |
||||
| 12 Foy BD, Rotem A, Toner M, Tompkins RG, Yarmush ML: A device to measure the oxygen uptake rate of attached cells: Importance in bioartificial organ design. Cell Transplant 1994;3:515-527. https://doi.org/10.1177/096368979400300609 |
||||
| 13 Giri S, Bader A: Improved preclinical safety assessment using micro-BAL devices: the potential impact on human discovery and drug attrition. Drug Discov Today 2011;16:382-397. https://doi.org/10.1016/j.drudis.2011.02.012 |
||||
| 14 Khetani SR, Bhatia SN: Micro scale culture of human liver cells for drug development. Nat Biotechnol 2008;26:120-126. https://doi.org/10.1038/nbt1361 |
||||
| 15 Gou M, Qu X, Zhu W, Xiang M, Yang J, Zhang K, Wei Y, Chen S: Bio-inspired detoxification using 3D-printed hydrogel Nano composites. Nat Commun 2014;5:3774. https://doi.org/10.1038/ncomms4774 |
||||
| 16 Materne EM, Tonevitsky AG, Marx U: Chip-based liver equivalents for toxicity testing--organotypicalness versus cost-efficient high throughput. Lab Chip 2013;13:3481-3495. https://doi.org/10.1039/c3lc50240f |
||||
| 17 Ebrahimkhani MR, Neiman JA, Raredon MS, Hughes DJ, Griffith LG: Bioreactor technologies to support liver function in vitro . Adv Drug Deliv Rev 2014;69-70:132-157. https://doi.org/10.1016/j.addr.2014.02.011 |
||||
| 18 Strassburg CP: Pharmacogenetics of Gilbert's syndrome. Pharmacogenomics 2008;9:703-715. https://doi.org/10.2217/14622416.9.6.703 |
||||
| 19 Herman B, Nieminen AL, Gores GJ, Lemasters JJ: Irreversible injury in anoxic hepatocytes precipitated by an abrupt increase in plasma membrane permeability. FASEB J 1988;2:146-151. https://doi.org/10.1096/fasebj.2.2.3342967 |
||||
| 20 Bader A, Fruhauf N, Tiedge M, Drinkgern M, De Bartolo L, Borlak JT, Steinhoff G, Haverich A: Enhanced oxygen delivery reverses anaerobic metabolic states in prolonged sandwich rat hepatocyte culture. Exp Cell Res 1999;246:221-232. https://doi.org/10.1006/excr.1998.4295 |
||||
| 21 Schmitmeier S, Langsch A, Jasmund I, Bader A: Development and characterization of a small-scale bioreactor based on a bioartificial hepatic culture model for predictive pharmacological in vitro screenings. Biotechnol Bioeng 2006;95:1198-1206. https://doi.org/10.1002/bit.21089 |
||||
| 22 Maringka M, Giri S, Bader A: Preclinical characterization of primary porcine hepatocytes in a clinically relevant flat membrane bioreactor. Biomaterials 2010;31:156-172. https://doi.org/10.1016/j.biomaterials.2009.09.020 |
||||
| 23 Fruhauf NR, Oldhafer KJ, Holtje M, Kaiser GM, Fruhauf JH, Stavrou GA, Bader A, Broelsch CE: A bioartificial liver support system using primary hepatocytes: a preclinical study in a new porcine hepatectomy model. Surgery 2004;136:47-56. https://doi.org/10.1016/j.surg.2003.12.017 |
||||
| 24 Bhatia SN, Ingber DE: Microfluidic organs-on-chips. Nat Biotechnol 2014;32:760-772. https://doi.org/10.1038/nbt.2989 |
||||
| 25 Strain AJ, Neuberger JM: A bioartificial liver--state of the art. Science 2002;295:1005-1009. https://doi.org/10.1126/science.1068660 |
||||
| 26 Stevens KM: Oxygen requirements for liver cells in vitro . Nature 1965;206:199. https://doi.org/10.1038/206199a0 |
||||
| 27 Jensen MD: Diffusion in tissue culture on gas-permeable and impermeable supports J. Theory Bio 1976;56:443-458. https://doi.org/10.1016/S0022-5193(76)80085-9 |
||||
| 28 Nahmias Y, Kramvis Y, Barbe L, Casali M, Berthiaume F, Yarmush ML: A novel formulation of oxygen-carrying matrix enhances liver-specific function of cultured hepatocytes. FASEB J 2006;20:2531-2533. https://doi.org/10.1096/fj.06-6192fje |
||||
| 29 Giri S, Weingartz U, Nieber K, Acikgoz A, Bader A: Cryopreservation of primary porcine liver cells in an organotypical sandwich model in a clinically relevant flat membrane bioreactor. Biotechnol Lett 2010;32:765-771. https://doi.org/10.1007/s10529-010-0228-6 |
||||
| 30 Gebhardt R, Mecke D: Perfused monolayer cultures of rat hepatocytes as an improved in vitro system for studies on ureogenesis. Exp Cell Res 1979;124:349-359. https://doi.org/10.1016/0014-4827(79)90210-6 |
||||
| 31 Dich J, Grunnet N: A perfusion system for cultured hepatocytes. Anal Biochem 1992;206:68-72. https://doi.org/10.1016/S0003-2697(05)80012-0 |
||||
| 32 Bai J, Rodriguez AM, Melendez JA, Cederbaum AI: Overexpression of catalase in cytosolic or mitochondrial compartment protects HepG2 cells against oxidative injury. J Biol Chem 1999;274:26217-26224. https://doi.org/10.1074/jbc.274.37.26217 |
||||
| 33 Pardo M, Tirosh O: Protective signaling effect of manganese superoxide dismutase in hypoxia-reoxygenation of hepatocytes. Free Radic Res 2009;43:1225-39. https://doi.org/10.3109/10715760903271256 |
||||
| 34 Zieve L, Shekleton M, Lyftogt C, Draves K: Ammonia, octanoate and a mercaptan depress regeneration of normal rat liver after partial hepatectomy. Hepatology 1985;5:28-31. https://doi.org/10.1002/hep.1840050107 |
||||
| 35 Bernal W, Hyyrylainen A, Gera A, Audimoolam VK, McPhail MJ, Auzinger G, Rela M, Heaton N, O'Grady JG, Wendon J, Williams R. Lessons from look-back in acute liver failure? A single center experience of 3300 patients. J Hepatol 2013;59:74-80. https://doi.org/10.1016/j.jhep.2013.02.010 |
||||